ប្រឆាំងនឹងភាពចាស់
Back Clinic Anti Aging Chiropractic and Functional Medicine Team ។ រាងកាយរបស់យើងស្ថិតនៅក្នុងសមរភូមិឥតឈប់ឈរ និងមិនចេះចប់ដើម្បីរស់រានមានជីវិត។ កោសិកាត្រូវបានចាប់កំណើត កោសិកាត្រូវបានបំផ្លាញ។ អ្នកវិទ្យាសាស្ត្រប៉ាន់ប្រមាណថាកោសិកានីមួយៗត្រូវតែទប់ទល់នឹងការវាយលុករបស់បុគ្គលលើសពី 10,000 ពីប្រភេទអុកស៊ីហ្សែនប្រតិកម្ម (ROS) ឬរ៉ាឌីកាល់សេរី។ បើគ្មានការបរាជ័យ រាងកាយមានប្រព័ន្ធព្យាបាលដោយខ្លួនឯងមិនគួរឱ្យជឿ ដែលទប់ទល់នឹងការវាយប្រហារ និងបង្កើតឡើងវិញនូវអ្វីដែលខូច ឬបំផ្លាញ។ នេះគឺជាភាពស្រស់ស្អាតនៃការរចនារបស់យើង។
ដើម្បីស្វែងយល់ពីជីវវិទ្យានៃភាពចាស់ និងបកប្រែការយល់ដឹងបែបវិទ្យាសាស្ត្រទៅជាអន្តរាគមន៍ដែលធ្វើអោយសុខភាពចុងដង្ហើមប្រសើរឡើងតាមរយៈការព្យាបាល។ វាមានប្រយោជន៍ក្នុងការមានទស្សនៈយល់ស្របច្បាស់លាស់អំពីអ្វីដែលពិតជាបង្កើតការព្យាបាលប្រឆាំងនឹងភាពចាស់។
តាំងពីមុនថ្ងៃនៃការស្វែងរកអាយុវែងរបស់ Ponce de Leon បុរសតែងតែត្រូវបានទាក់ទាញដោយឱកាសនៃយុវវ័យដ៏អស់កល្បជានិច្ច។ ការថែទាំតាមបែប Chiropractic ជាមួយនឹងចលនាសុខភាពរបស់វាគឺជាវិធីសាស្រ្តដ៏មានអានុភាពនៃស្ថេរភាព និងបង្កើនសមត្ថភាពព្យាបាលដោយខ្លួនឯងនេះ។ វេជ្ជបណ្ឌិត Alex Jimenez ពិភាក្សាអំពីគោលគំនិតជុំវិញការប្រឆាំងភាពចាស់ pandora ។
.
by វេជ្ជបណ្ឌិត Alex Jimenez ការព្យាបាលដោយចាក់ម្ជុលវិទ្យាសាស្ត្រ , ប្រឆាំងនឹងភាពចាស់
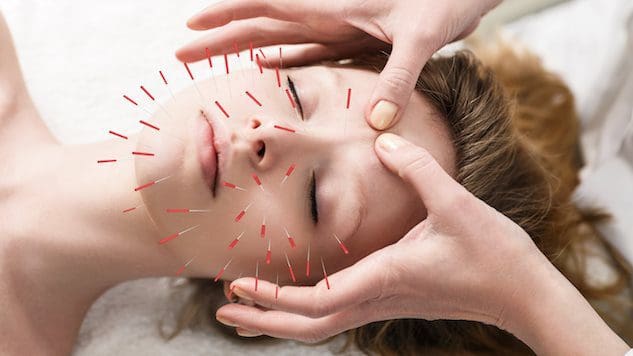
សម្រាប់បុគ្គលដែលចង់កែលម្អ ឬថែរក្សាសុខភាពស្បែក តើការចាក់ម្ជុលវិទ្យាសាស្ត្រអាចជួយកែលម្អស្បែក និងប្រឆាំងនឹងដំណើរការនៃភាពចាស់បានទេ?
ការចាក់ម្ជុលវិទ្យាសាស្ត្រ
ការចាក់ម្ជុលវិទ្យាសាស្ត្រតាមបែបប្រពៃណីនៃការចាក់ម្ជុលវិទ្យាសាស្ត្រ។ គោលបំណងគឺដើម្បីបញ្ច្រាសសញ្ញានៃភាពចាស់ និងធ្វើឱ្យសុខភាពស្បែកប្រសើរឡើង។ ជួនកាលវាត្រូវបានគេហៅថាការចាក់ម្ជុលវិទ្យាសាស្ត្រឡើងវិញ ដែលត្រូវបានគេប្រើជាជម្រើសមួយសម្រាប់ការលើកមុខវះកាត់ និងនីតិវិធីសាមញ្ញផ្សេងទៀត។ ការសិក្សាបឋមបានពិនិត្យមើលពីរបៀបដែលវាអាចជួយលុបបំបាត់ស្នាមជ្រួញតាមអាយុ លើកត្របកភ្នែកស្រវាំង និងកាត់បន្ថយស្នាមជ្រួញ។ (Younghee Yun et al., 2013 )
របៀបដែលការចាក់ម្ជុលវិទ្យាសាស្ត្រដំណើរការ
នៅក្នុងឱសថបុរាណចិន ឬ TCM ការចាក់ម្ជុលវិទ្យាសាស្ត្រត្រូវបានប្រើប្រាស់ជាយូរមកហើយដើម្បីកែលម្អលំហូរថាមពល - qi ឬ chi - នៅទូទាំងរាងកាយ។ ថាមពលនេះត្រូវបានគេជឿថាចរាចរតាមរយៈផ្លូវថាមពលដែលគេស្គាល់ថាជា meridians ។ នៅពេលដែលបញ្ហាសុខភាពកើតឡើងយោងទៅតាម TCM មានការស្ទះឬការស្ទះនៅក្នុងឈាមរត់។វិទ្យាស្ថានជាតិសុខាភិបាលឆ្នាំ ២០១៧ )
ការចាក់ម្ជុលវិទ្យាសាស្ត្រ
ការចាក់ម្ជុលវិទ្យាសាស្ត្រត្រូវបានគេនិយាយថាធ្វើឱ្យប្រសើរឡើងនូវសុខភាពស្បែក និងដើរតួជាការព្យាបាលប្រឆាំងនឹងភាពចាស់ដោយការជំរុញការផលិតកូឡាជែន។ ប្រូតេអ៊ីននេះគឺជាសមាសធាតុសំខាន់នៃស្បែក។ ស្រទាប់ខាងក្នុងនៃស្បែកបាត់បង់ កូឡាជែន និងភាពរឹងមាំនៅពេលដែលរាងកាយកាន់តែចាស់។ ទោះជាយ៉ាងណាក៏ដោយ ការស្រាវជ្រាវបន្ថែមគឺត្រូវការជាចាំបាច់ដើម្បីគាំទ្រការអះអាងដែលថាការចាក់ម្ជុលវិទ្យាសាស្ត្រអាចជំរុញការផលិតកូឡាជែន។ អ្នកខ្លះណែនាំថាការចាក់ម្ជុលវិទ្យាសាស្ត្រជួយឱ្យស្បែកមានភាពក្មេងជាងវ័យដោយការធ្វើឱ្យប្រសើរឡើងនូវថាមពលរួមរបស់រាងកាយ។ ការសិក្សាមួយបានរកឃើញថា បុគ្គលបានឃើញភាពប្រសើរឡើងបន្ទាប់ពីវគ្គ 5 នៃការចាក់ម្ជុលវិទ្យាសាស្ត្រលើផ្ទៃមុខ។ (Younghee Yun et al., 2013 ) ទោះបីជាយ៉ាងណាក៏ដោយ វាត្រូវបានណែនាំអោយធ្វើការព្យាបាលចំនួន 10 ម្តង ឬ 2 ដងក្នុងមួយសប្តាហ៍ដើម្បីទទួលបានលទ្ធផលល្អបំផុត។ បន្ទាប់ពីនោះ ការព្យាបាលថែទាំត្រូវបានធ្វើឡើងរៀងរាល់ 4 ទៅ 8 សប្តាហ៍។ មិនដូច Botox ឬ dermal fillers ការចាក់ម្ជុលវិទ្យាសាស្ត្រមិនមែនជាការជួសជុលរហ័សនោះទេ។ ការផ្តោតអារម្មណ៍គឺដើម្បីបង្កើតការផ្លាស់ប្តូររយៈពេលវែងនៃស្បែក និងរាងកាយ ដែលមានន័យថាមានភាពប្រសើរឡើង៖
នៅពេលដែលម្ជុលត្រូវបានបញ្ចូលទៅក្នុងស្បែក ពួកគេបង្កើតរបួសដែលគេស្គាល់ថាជា microtraumas វិជ្ជមាន។ សមត្ថភាពព្យាបាល និងជួសជុលធម្មជាតិរបស់រាងកាយនឹងមានសកម្មភាពនៅពេលវាដឹងរបួសទាំងនេះ។ ស្នាមប្រេះទាំងនេះជំរុញប្រព័ន្ធឡាំហ្វាទិច និងឈាមរត់ ដែលបញ្ជូនសារធាតុចិញ្ចឹម និងអុកស៊ីហ្សែនទៅកោសិកាស្បែក ចិញ្ចឹមពួកវាពីខាងក្នុងចេញមកក្រៅ។
នេះជួយឱ្យស្បែកមុខរលោង និងជួយឱ្យស្បែកភ្លឺថ្លា។
microtraumas វិជ្ជមានក៏ជំរុញការផលិតកូឡាជែនផងដែរ។
នេះជួយកែលម្អភាពបត់បែន កាត់បន្ថយបន្ទាត់ និងស្នាមជ្រួញ។
ជម្មើសជំនួស
ឱសថធម្មជាតិជាច្រើនអាចជួយកែលម្អសុខភាពស្បែក និងផ្តល់អត្ថប្រយោជន៍ប្រឆាំងនឹងភាពចាស់។ Ceramides គឺជាម៉ូលេគុលជាតិខ្លាញ់ដែលត្រូវបានរកឃើញដោយធម្មជាតិនៅក្នុងស្រទាប់ខាងលើនៃស្បែក និងជាធាតុផ្សំដែលប្រើក្នុងផលិតផលថែរក្សាស្បែក។ ទាំងនេះអាចការពារប្រឆាំងនឹងភាពស្ងួតដែលទាក់ទងនឹងភាពចាស់នៃស្បែក។ (L Di Marzio ឆ្នាំ ២០០៨ ការស្រាវជ្រាវបឋមបានបង្ហាញថា ការលាបតែសលើស្បែកអាចប្រឆាំងនឹងការបំបែកនៃ collagen និង elastin ដែលជាប្រូតេអ៊ីនដែលជួយដល់ការបត់បែនស្បែក និងការពារការយារធ្លាក់)។ វាក៏មានភស្តុតាងដែលថាសារធាតុធម្មជាតិដូចជាប្រេង argan ប្រេង borage និងសមុទ្រ buckthorn អាចផ្តល់អត្ថប្រយោជន៍សំណើមដែលអាចធ្វើអោយស្បែកប្រសើរឡើង។(Tamsyn SA Thring et al., 2009 )
ខណៈពេលដែលភស្តុតាងបន្ថែមទៀតនៃការចាក់ម្ជុលវិទ្យាសាស្ត្រគឺត្រូវការជាចាំបាច់ ការបញ្ចូលការចាក់ម្ជុលវិទ្យាសាស្ត្រអាចជួយគ្រប់គ្រងភាពតានតឹង និងពង្រឹងសុខភាពទូទៅ។ បុគ្គលដែលពិចារណាលើការចាក់ម្ជុលវិទ្យាសាស្ត្រគួរទៅពិគ្រោះជាមួយអ្នកផ្តល់សេវាថែទាំសុខភាពបឋមរបស់ពួកគេដើម្បីមើលថាតើវាត្រឹមត្រូវសម្រាប់ពួកគេឬអត់។
ការលើកកម្ពស់សុខភាពរួមគ្នា៖ ការទទួលយកការវាយតម្លៃ និងការព្យាបាលពហុជំនាញ
VIDEO
ឯកសារយោង
Yun, Y., Kim, S., Kim, M., Kim, K., Park, JS, & Choi, I. (2013) ។ ឥទ្ធិពលនៃការចាក់ម្ជុលវិទ្យាសាស្ត្រលើផ្ទៃមុខ លើភាពយឺតនៃផ្ទៃមុខ៖ ការសិក្សាសាកល្បងដៃតែមួយ។ ការបំពេញបន្ថែម និងថ្នាំជំនួសដោយផ្អែកលើភស្តុតាង៖ eCAM, 2013, 424313។ doi.org/10.1155/2013/424313
មជ្ឈមណ្ឌលជាតិសម្រាប់ការបំពេញបន្ថែម និងឱសថជំនួស។ (២០០៧)។ ការចាក់ម្ជុលវិទ្យាសាស្ត្រ៖ ការណែនាំ។ គេហទំព័រមជ្ឈមណ្ឌលជាតិសម្រាប់បំពេញបន្ថែម និងឱសថជំនួស។ choimd.com/downloads/NIH-info-on-acupuncture.pdf
Kuge, H., Mori, H., Tanaka, TH, & Tsuji, R. (2021) ។ ភាពជឿជាក់ និងសុពលភាពនៃសន្លឹកពិនិត្យផ្ទៃមុខ (FCS)៖ បញ្ជីពិនិត្យសម្រាប់ការពេញចិត្តដោយខ្លួនឯងជាមួយនឹងការចាក់ម្ជុលវិទ្យាសាស្ត្រ។ ឱសថ (Basel, Switzerland), ៨(៤), ១៨. doi.org/10.3390/medicines8040018
Di Marzio, L., Cinque, B., Cupelli, F., De Simone, C., Cifone, MG, & Giuliani, M. (2008) ។ ការកើនឡើងនៃកម្រិតសេរ៉ាមិចស្បែកចំពោះមនុស្សវ័យចំណាស់បន្ទាប់ពីការប្រើប្រាស់ថ្នាំ sphingomyelinase បាក់តេរីក្នុងរយៈពេលខ្លីពី Streptococcus thermophilus ។ ទិនានុប្បវត្តិអន្តរជាតិនៃ immunopathology និងឱសថវិទ្យា, 21(1), 137–143 ។ doi.org/10.1177/039463200802100115
Thring, TS, Hili, P., & Naughton, DP (2009) ។ Anti-collagenase, anti-elastase និងសកម្មភាពប្រឆាំងអុកស៊ីតកម្មនៃសារធាតុចម្រាញ់ពីរុក្ខជាតិចំនួន 21 ។ ការបំពេញបន្ថែម និងថ្នាំជំនួស BMC, 9, 27 ។ doi.org/10.1186/1472-6882-9-27
by វេជ្ជបណ្ឌិត Alex Jimenez ប្រឆាំងនឹងភាពចាស់
រក្សាឆ្អឹងខ្នងរបស់មនុស្សម្នាក់ឱ្យស្ថិតក្នុងទម្រង់កំពូលស្មើនឹងការឈឺចាប់តិចនិងភាពចល័តភាពបត់បែននិងសេរីភាពកាន់តែច្រើន ។ រាងកាយចុះខ្សោយហើយជាឥទ្ធិពលធម្មជាតិនៃភាពចាស់ដែលកើតឡើងចំពោះយើងគ្រប់គ្នា។ បញ្ហាឆ្អឹងខ្នងទាក់ទងនឹងភាពចាស់អាចក្លាយជារឿងធ្ងន់ធ្ងរប្រសិនបើមិនត្រូវបានដោះស្រាយនិងអនុម័តលើការធ្វើលំហាត់ប្រាណការលាតសន្ធឹងនិងការថែទាំតាមបែបជីវ។
ភាពចាស់ជរានិងខ្នង
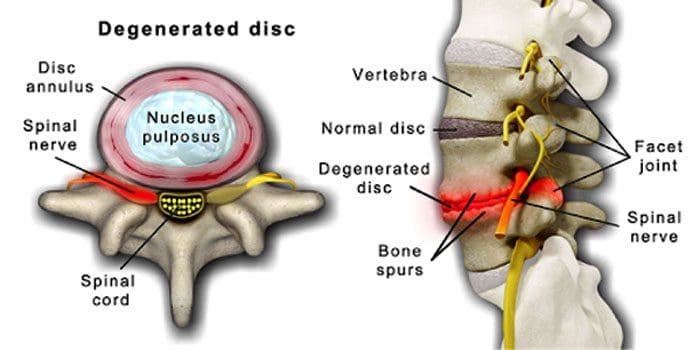
វាជារឿងធម្មតាទេដែលឌីសឆ្អឹងខ្នងនិងសន្លាក់កាន់តែយ៉ាប់យ៉ឺនទៅតាមអាយុ ។ ជម្ងឺក្រិនឆ្អឹងខ្នងឬការរួមតូចនៃប្រឡាយឆ្អឹងខ្នងក៏អាចជាផ្នែកមួយនៃដំណើរការចាស់ដែរ។ លក្ខខណ្ឌពីរដែលបានកើតឡើងដោយសារភាពចាស់ជរា ជំងឺឌីសខូច និង ជំងឺរលាកសន្លាក់ ដែលអាចរួមបញ្ចូលផងដែរ ការរឹងនៃសរសៃចងឆ្អឹងខ្នងនិងជំងឺពុកឆ្អឹង .
ជំងឺឌីសឌីសត្រូវបានជួបប្រទះដោយបុគ្គល 40% នៃអាយុ 40 ឆ្នាំ
កើនឡើងដល់ ៨០% សម្រាប់បុគ្គលដែលមានអាយុចាប់ពី ៨០ ឆ្នាំឡើងទៅ។
វាផ្តោតជុំវិញ ឌីសដែលផ្លាស់ប្តូរបន្តិចម្តង ៗ ពីភាគច្រើនទៅជាជាតិខ្លាញ់ភាគច្រើន .
នៅពេលវាធាត់ឌីសប្រែជាតូចចង្អៀតនិងបាត់បង់ការបត់បែន។
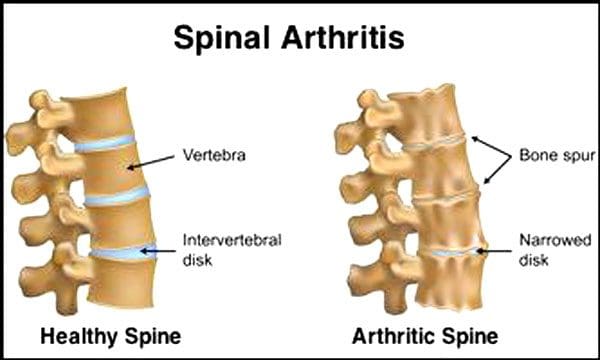
មជ្ឍមណ្ឌលត្រួតពិនិត្យនិងបង្ការជំងឺនិយាយថា ២៣% នៃមនុស្សពេញវ័យជនជាតិអាមេរិកមានជំងឺរលាកសន្លាក់ ។ នេះគឺជាលក្ខខណ្ឌមួយដែលជះឥទ្ធិពលជាចម្បងលើសន្លាក់ facet ។ សន្លាក់ប្រែជាហើមដែលជួយកាត់បន្ថយចលនានិងអាចប៉ះពាល់ដល់សរសៃប្រសាទឆ្អឹងខ្នងដែលបណ្តាលឱ្យឈឺចាប់ខ្សោយនិងខ្សោយ។ ជាមួយនឹងពេលវេលាសរសៃនៅជុំវិញនិងក្នុងឆ្អឹងខ្នងរឹងកាត់បន្ថយជួរនៃចលនាបណ្តាលឱ្យមានស្នាមប្រឡាក់ ។ ការបាត់បង់ឆ្អឹងឬជំងឺពុកឆ្អឹងត្រូវបាននាំមកដោយការផ្លាស់ប្តូរអ័រម៉ូននិងកត្តាដទៃទៀតដូចជាអាហារូបត្ថម្ភ។ ភាពចាស់ជរាគឺជាដំណើរការធម្មជាតិមួយប៉ុន្តែបុគ្គលម្នាក់ៗអាចជួយឱ្យឆ្អឹងខ្នងរបស់ពួកគេស្ថិតក្នុងទម្រង់កំពូលមិនថាពួកគេមានអាយុប៉ុន្មាននោះទេ។
ការអនុវត្តឥរិយាបថសុខភាព
បិទដំបង មេកានិចរាងកាយដែលមានសុខភាពល្អត្រឹមត្រូវ
ខួរឆ្អឹងខ្នង
ជំងឺឌីសស្តារ
Herniation
ហានិភ័យនៃការបាក់ឆ្អឹងឆ្អឹងខ្នង
ការអនុវត្តឥរិយាបថត្រឹមត្រូវរួមមាន៖
កាត់បន្ថយការរអិល
ត្រូវប្រាកដថាស្ថានីយ៍ការងារមានទំរង់ខាងលើនិងមានសម្លេងល្អ
សកម្មភាពណាក៏ដោយដែលបុគ្គលម្នាក់ៗចូលរួមត្រូវព្យាយាមធ្វើ ពន្លូតនិងធ្វើឱ្យឆ្អឹងខ្នងវែង
វិធីសាស្រ្តនេះក៏អនុវត្តផងដែរដើម្បីលើក។
ត្រូវប្រាកដថាពត់ជង្គង់នៅពេលលើកនិងរក្សាឆ្អឹងខ្នងបញ្ឈរតាមដែលអាចធ្វើទៅបាន។
ហា្កបាន
ហា្កបាន អាចមានអត្ថប្រយោជន៍ខ្ពស់សម្រាប់ឆ្អឹងខ្នងដែលមានសុខភាពល្អនិងក្មេងជាងវ័យ។ យូហ្គាបំពេញផ្នែកបីសម្រាប់រក្សាឆ្អឹងខ្នងក្នុងទម្រង់ខាងលើ ។ នេះរួមមានៈ
លំហាត់ជាប្រចាំ
រក្សាភាពបត់បែន
សម្រេចបាននូវទំងន់រាងកាយល្អ
យូហ្គាគឺជាសកម្មភាពដែលអាចការពារអាយុសម្រាប់ឆ្អឹងខ្នង។ ពីព្រោះវា:
រក្សាភាពរឹងមាំ
ភាពបត់បែន
រូបភាព
តុល្យភាព
អាចមានប្រយោជន៍សម្រាប់ភាពខុសគ្នានៃស្ថានភាពឆ្អឹងខ្នងជាពិសេសការឈឺចាប់រលាកសន្លាក់
ការដួលអាចបណ្តាលឱ្យរងរបួសធ្ងន់ធ្ងរ។ យូហ្គាក៏អាចជួយធ្វើឱ្យមានតុល្យភាពផងដែរ។
មើល Chiropractor
ថ្នាំបង្ការគឺជាគន្លឹះក្នុងការធ្វើឱ្យរាងកាយមានសុខភាពល្អវ័យក្មេងនិងរឹងមាំតាមដែលអាចធ្វើទៅបាន។ ការពិនិត្យលើប្រដាប់បន្តពូជអាចកំណត់ថាតើមានបញ្ហាឆ្អឹងខ្នងនិងការធ្វើរោគវិនិច្ឆ័យដើម្បីបង្កើតផែនការព្យាបាលមួយដែលប្រសើរបំផុតឬអត់។ ប្រសិនបើមុខងាររាងកាយមានកំណត់ដោយសារតែការឈឺចាប់នៅខាងក្រោយនិង / ឬជើង។ ទាក់ទងការរងរបួសផ្នែកវេជ្ជសាស្ត្រផ្នែកជីវសាស្រ្តនិងគ្លីនិកវេជ្ជសាស្ត្រដែលមានមុខងារនិងយកឆ្អឹងខ្នងត្រឡប់មកវិញតាមទម្រង់ខាងលើ
សមាសភាពរាងកាយ
VIDEO
លំហាត់គ្រាប់បាល់ / ស្ថេរភាព
លំហាត់នេះ ធ្វើការជាក្រុមសាច់ដុំជាក់លាក់ចំពោះកម្លាំងឆ្អឹងខ្នងនិងរួមមាន៖
សរសៃពួរគន្លាក់ជើង
រីករាយ
ពោះជ្រៅ
អ្នកចាប់ពង្រត់និងបង្វិលត្រគាក
ការធ្វើលំហាត់ប្រាណបែបនេះគឺជាវិធីមួយដែលមានប្រសិទ្ធភាពបំផុតក្នុងការបង្កើតកម្លាំងមុខងារនិងការស៊ូទ្រាំនៅត្រង់ត្រគាកត្រគាកនិងការពារការរងរបួស។ ដើម្បីធ្វើការសាកល្បងនេះ៖
កុហកលើខ្នងរបស់អ្នកដោយជង្គង់បត់
លើកជើងឡើងលើដូច្នេះបាតជើងស្ថិតនៅលើកំពូលនៃបាល់ហាត់ប្រាណ
រមៀលជើងរបស់អ្នកចេញរហូតដល់ពួកគេត្រង់
កាន់ជំហរមួយវិនាទីឬពីរ
ត្រឡប់ទៅផ្នែកខាងលើនៃចលនាខណៈពេលដែលច្របាច់សរសៃពួរ
ធ្វើការសាច់ដុំទាំងនេះនឹងជួយធ្វើឱ្យចលនាញាប់ញ័រសួតឬពត់កោងងាយស្រួលនៅលើឆ្អឹងខ្នង។
វេជ្ជបណ្ឌិតអាឡិចហ្សង់ហ្សេនហ្សេសមិនទទួលបន្ទុកប្រកាសប្លក់
វិសាលភាពនៃព័ត៌មានរបស់យើងត្រូវបានកំណត់ចំពោះរោគសាស្ត្រសាច់ដុំឆ្អឹងសាច់ដុំរាងកាយសុខភាពនិងបញ្ហាសុខភាពរសើបនិង / ឬអត្ថបទវេជ្ជសាស្ត្រមុខងារប្រធានបទនិងការពិភាក្សា។ យើងប្រើពិធីសារសុខភាពនិងសុខភាពដែលមានមុខងារដើម្បីព្យាបាលនិងគាំទ្រដល់ការរងរបួសឬការរំខាននៃប្រព័ន្ធសាច់ដុំ។ ប្រកាសប្រធានបទប្រធានបទនិងការយល់ដឹងរបស់យើងទាក់ទងនឹងបញ្ហាគ្លីនិកបញ្ហានិងប្រធានបទដែលទាក់ទងនិងគាំទ្រដោយផ្ទាល់ឬដោយប្រយោលនូវវិសាលភាពនៃការអនុវត្តជាក់ស្តែងរបស់យើង។ *
ការិយាល័យរបស់យើងបានខិតខំប្រឹងប្រែងសមហេតុផលដើម្បីផ្តល់នូវឯកសារយោងដែលមានការគាំទ្រនិងបានកំណត់ការសិក្សាស្រាវជ្រាវដែលពាក់ព័ន្ធឬការសិក្សាគាំទ្រដល់ប្រកាសរបស់យើង។ យើងក៏ធ្វើការថតចម្លងការសិក្សាស្រាវជ្រាវដែលមានដល់ក្រុមប្រឹក្សាភិបាលឬសាធារណៈជនតាមការស្នើសុំ។ យើងយល់ថាយើងគ្របដណ្តប់លើបញ្ហានានាដែលត្រូវការការពន្យល់បន្ថែមអំពីវិធីដែលវាអាចជួយក្នុងផែនការថែទាំជាក់លាក់ឬពិធីសារព្យាបាល។ ដូច្នេះដើម្បីពិភាក្សាបន្ថែមលើប្រធានបទខាងលើសូមមេត្តាសួរលោកបណ្ឌិត Alex Jimenez ឬទំនាក់ទំនងមកលេខ ៩១៥-៨៥០-០៩០០ ។ អ្នកផ្តល់សេវាមានអាជ្ញាប័ណ្ណនៅរដ្ឋតិចសាស់និងញូម៉ិកស៊ិក *
ឯកសារយោង
សេចក្តីណែនាំ៖ � ការវាយតំលៃលើបច្ចេកវិទ្យាសុខភាព Ontario �ស៊េរី។ �(ខែមេសា 2006) �ឌីសសិប្បនិម្មិតសម្រាប់ជំងឺឌីសឆ្អឹងកងខ្នង និងមាត់ស្បូន -ធ្វើបច្ចុប្បន្នភាព៖ ការវិភាគផ្អែកលើភស្តុតាង��pubmed.ncbi.nlm.nih.gov/23074480/
សេចក្តីណែនាំ៖ � មជ្ឈមណ្ឌលគ្រប់គ្រងនិងបង្ការជំងឺ។ �(ខែវិច្ឆិកា ២០២០) �ជំងឺរលាកសន្លាក់��www.cdc.gov/chronicdisease/resources/publications/factsheets/arthritis.htm
by វេជ្ជបណ្ឌិត Alex Jimenez ប្រឆាំងនឹងភាពចាស់ , របបអាហារ , Epigenetic របស់ , ឱសថមុខងារ , សុខភាព , សុខភាពធម្មជាតិ , អាហាររូបត្ថម្ភ , ជីវជាតិចំណីអាហារ , សុខុមាលភាព
អាហារដែលយើងញ៉ាំអាចមានសក្តានុពលដែលអាចផ្តល់អត្ថប្រយោជន៍ឬបង្កគ្រោះថ្នាក់ដល់សុខភាពរបស់យើង។ អាហាររូបត្ថម្ភខ្សោយអាចបណ្តាលឱ្យមានបញ្ហាសុខភាពជាច្រើនរួមមានភាពធាត់ជំងឺបេះដូងនិងជំងឺទឹកនោមផ្អែមប្រភេទទី ២ ។ ទន្ទឹមនឹងនេះសារធាតុចិញ្ចឹមត្រឹមត្រូវអាចធ្វើឱ្យអ្នកមានអារម្មណ៍ស្វាហាប់កាត់បន្ថយហានិភ័យនៃបញ្ហាសុខភាពក៏ដូចជាជួយរក្សានិងរក្សាទម្ងន់ឱ្យមានសុខភាពល្អ។ ប្រសិនបើអ្នកចង់លើកកម្ពស់ភាពជាប់បានយូរអ្នកត្រូវដុតរាងកាយរបស់អ្នកដោយអាហារល្អ ៗ ។ នៅក្នុងអត្ថបទបន្ទាប់យើងនឹងរាយបញ្ជីអាហារល្អ ៗ ជាច្រើនដែលទីបំផុតអាចជួយលើកកម្ពស់អាយុយឺនយូរដោយជួយលើកកម្ពស់សុខភាពនិងសុខភាពទូទៅផងដែរ។
បន្លែគួរឱ្យស្អប់ខ្ពើម
បន្លែដ៏គួរឱ្យស្អប់ខ្ពើមមានសមត្ថភាពពិសេសក្នុងការផ្លាស់ប្តូរអរម៉ូនរបស់យើងកេះប្រព័ន្ធរាងកាយបំបាត់ជាតិពុលធម្មជាតិហើយថែមទាំងជួយកាត់បន្ថយការលូតលាស់កោសិកាមហារីកទៀតផង។ ទាំងនេះត្រូវតែត្រូវបានទំពារយ៉ាងហ្មត់ចត់ឬញ៉ាំខ្ទិះខ្ទិះលាយឬលាយចូលគ្នាដើម្បីបញ្ចេញនូវគុណប្រយោជន៍របស់វា។ ស៊ុលហ្វួរ៉ាហ្វានដែលត្រូវបានគេរកឃើញនៅក្នុងបន្លែដែលត្រូវបានគេឆ្កាងក៏ត្រូវបានគេរកឃើញផងដែរដើម្បីជួយការពារជញ្ជាំងសរសៃឈាមពីការរលាកដែលអាចបង្កឱ្យមានជំងឺបេះដូង។ បន្លែដ៏គួរឱ្យស្អប់ខ្ពើមដូចជាខាត់ណាស្ពៃក្តោបពន្លកស៊ែលផ្កាខាត់ណាខៀវនិងផ្កាខាត់ណាខៀវគឺជាអាហារដែលសំបូរទៅដោយសារធាតុចិញ្ចឹមដែលក្រាស់បំផុតនៅក្នុងពិភពលោក។
សាឡាត់បៃតង
បន្លែស្លឹកបៃតងឆៅមានតិចជាង ១០០ កាឡូរីក្នុងមួយផោនដែលធ្វើឱ្យពួកគេក្លាយជាអាហារដ៏ល្អឥតខ្ចោះសម្រាប់ការសម្រកទម្ងន់។ ការទទួលទានបន្លែសាឡាដបន្ថែមទៀតក៏ត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការថយចុះហានិភ័យនៃការគាំងបេះដូងជំងឺដាច់សរសៃឈាមខួរក្បាលជំងឺទឹកនោមផ្អែមនិងជំងឺមហារីកជាច្រើនប្រភេទផងដែរ។ បន្លែស្លឹកបៃតងឆៅក៏សំបូរទៅដោយជីវជាតិ B-Vitamin folate សំខាន់ៗផងដែររួមទាំងសារជាតិ lutein និង zeaxanthin, carotenoids ដែលអាចជួយការពារភ្នែក។ សារធាតុគីមីដែលរលាយក្នុងខ្លាញ់ដូចជាការ៉ូណូទីនត្រូវបានគេរកឃើញនៅក្នុងបន្លែសាឡាត់ដូចជាសាឡាត់ស្ពៃខ្មៅស្ពៃបៃតងនិងស្លឹកស្ពៃបៃតងក៏មានឥទ្ធិពលប្រឆាំងអុកស៊ីតកម្មនិងប្រឆាំងនឹងការរលាកនៅក្នុងរាងកាយផងដែរ។
គ្រាប់
គ្រាប់គឺជាអាហារដែលមានជាតិ glycemic ទាបនិងជាប្រភពដ៏ល្អនៃខ្លាញ់ដែលមានសុខភាពល្អប្រូតេអ៊ីនរុក្ខជាតិជាតិសរសៃសារធាតុប្រឆាំងអុកស៊ីតកម្ម phytosterols និងសារធាតុរ៉ែដែលជួយកាត់បន្ថយបន្ទុកគ្លីសេរីននៃអាហារទាំងមូលដែលធ្វើឱ្យវាក្លាយជាផ្នែកមួយដ៏សំខាន់នៃការប្រឆាំងនឹងជំងឺទឹកនោមផ្អែម។ របបអាហារ ដោយមិនគិតពីដង់ស៊ីតេកាឡូរីរបស់ពួកគេការញ៉ាំគ្រាប់ផ្លែឈើអាចជួយលើកកម្ពស់ការសម្រកទម្ងន់។ គ្រាប់ក៏អាចជួយកាត់បន្ថយកូលេស្តេរ៉ុលនិងជួយកាត់បន្ថយហានិភ័យនៃជំងឺបេះដូង។
គ្រាប់
គ្រាប់ជាច្រើនដូចជាគ្រាប់ក៏ផ្តល់នូវជាតិខ្លាញ់ដែលមានសុខភាពល្អអង់ទីអុកស៊ីដង់និងសារធាតុរ៉ែផងដែរទោះយ៉ាងណាវាមានជាតិប្រូតេអ៊ីនច្រើនហើយសំបូរទៅដោយជាតិរ៉ែរ៉ែ។ ជីឆៃ flax និងគ្រាប់ hemp សំបូរទៅដោយខ្លាញ់អូមេហ្គា ៣ ។ គ្រាប់ជីឆៃ flax និងគ្រាប់ល្ងក៏ជាលីនថេនសំបូរឬ phytoestrogens ប្រឆាំងនឹងជំងឺមហារីកសុដន់ផងដែរ។ ម៉្យាងទៀតគ្រាប់ល្ងសំបូរទៅដោយជាតិកាល់ស្យូមនិងវីតាមីនអ៊ីហើយគ្រាប់ល្ពៅសំបូរទៅដោយជាតិស័ង្កសី។
Berries
ផ្លែប៊ឺរីគឺជាផ្លែឈើដែលមានសារធាតុប្រឆាំងអុកស៊ីតកម្មដែលអាចជួយលើកកម្ពស់សុខភាពបេះដូង។ ការសិក្សាស្រាវជ្រាវដែលអ្នកចូលរួមបរិភោគផ្លែស្ត្របឺរីឬប៊្លូបឺរីជារៀងរាល់ថ្ងៃអស់រយៈពេលជាច្រើនសប្តាហ៍បានរាយការណ៍ថាមានការកើនឡើងសម្ពាធឈាមសរុបនិងកូឡេស្តេរ៉ុលអាក្រក់អិល។ ផ្លែប៊ឺរីក៏មានមុខងារប្រឆាំងនឹងជំងឺមហារីកហើយត្រូវបានបង្ហាញថាជួយការពារការធ្លាក់ចុះនៃការយល់ដឹងដែលទាក់ទងនឹងភាពចាស់។
ផ្លែទទឹម
សារធាតុគីមីដែលត្រូវបានគេស្គាល់ច្រើនជាងគេបំផុតនៅក្នុងផ្លែទទឹមគឺភូទីហ្គីនទទួលខុសត្រូវចំពោះសកម្មភាពអុកស៊ីតកម្មរបស់ផ្លែឈើជាងពាក់កណ្តាល។ សារធាតុគីមីទទឹមមានសារធាតុប្រឆាំងនឹងជំងឺមហារីកការការពារបេះដូងនិងសុខភាពខួរក្បាល។ នៅក្នុងការសិក្សាស្រាវជ្រាវមួយមនុស្សវ័យចំណាស់ដែលផឹកទឹកផ្លែទទឹមជារៀងរាល់ថ្ងៃរយៈពេល ២៨ ថ្ងៃបានធ្វើតេស្តិ៍ការចងចាំបានល្អប្រសើរបើប្រៀបធៀបទៅនឹងអ្នកដែលទទួលទានភេសជ្ជៈ placebo ។
សណ្តែក
ការបរិភោគសណ្តែកនិងសណ្តែកដទៃទៀតអាចជួយធ្វើឱ្យមានតុល្យភាពជាតិស្ករក្នុងឈាមកាត់បន្ថយចំណង់អាហារនិងការពារប្រឆាំងនឹងជំងឺមហារីកពោះវៀនធំ។ សណ្តែកគឺជាអាហារប្រឆាំងនឹងជំងឺទឹកនោមផ្អែមដែលអាចជួយលើកកម្ពស់ការសម្រកទម្ងន់ព្រោះវាត្រូវបានរំលាយយឺត ៗ ដែលធ្វើឱ្យកម្រិតជាតិស្ករក្នុងឈាមថយចុះបន្ទាប់ពីបរិភោគហើយជួយការពារការឃ្លានអាហារដោយជំរុញការឆ្អែត។ ការបរិភោគសណ្តែកនិងសណ្តែកដទៃទៀតពីរដងក្នុងមួយសប្តាហ៍ត្រូវបានគេរកឃើញថាអាចបន្ថយហានិភ័យនៃជំងឺមហារីកពោះវៀនធំ។ ការបរិភោគសណ្តែកនិងសណ្តែកដទៃទៀតដូចជាសណ្តែកក្រហមសណ្តែកខ្មៅម្ទេសសណ្តែកនិងសណ្តែកពកក៏ផ្តល់នូវការការពារគួរឱ្យកត់សម្គាល់ប្រឆាំងនឹងជំងឺមហារីកដទៃទៀតដែរ។
ផ្សិត
ការបរិភោគផ្សិតជាប្រចាំត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការថយចុះហានិភ័យនៃជំងឺមហារីកសុដន់។ ផ្សិតសនិងផូបាឡូឡូមានអត្ថប្រយោជន៍ជាពិសេសប្រឆាំងនឹងជំងឺមហារីកសុដន់ព្រោះវាមានសារធាតុ aromatase inhibitors ឬសមាសធាតុផ្សំដែលរារាំងដល់ការផលិតអរម៉ូនអ៊ឹស្ត្រូសែន។ ផ្សិតបានបង្ហាញថាមានឥទ្ធិពលប្រឆាំងនឹងការរលាកក៏ដូចជាផ្តល់នូវសកម្មភាពកោសិកាភាពស៊ាំកាន់តែប្រសើរឡើងការការពារការបំផ្លាញឌីអិនអេការថយចុះការលូតលាស់កោសិកាមហារីកនិងការរារាំង angiogenesis ។ ផ្សិតគួរតែត្រូវបានចម្អិនជានិច្ចព្រោះផ្សិតឆៅមានសារធាតុគីមីបង្កមហារីកដែលត្រូវបានគេស្គាល់ថា agaritine ដែលត្រូវបានកាត់បន្ថយយ៉ាងខ្លាំងដោយការចម្អិនអាហារ។
ខ្ទឹមបារាំងនិងខ្ទឹមស
ខ្ទឹមបារាំងនិងខ្ទឹមសផ្តល់អត្ថប្រយោជន៍ដល់ប្រព័ន្ធសរសៃឈាមបេះដូងនិងប្រព័ន្ធភាពស៊ាំក៏ដូចជាផ្តល់នូវឥទ្ធិពលប្រឆាំងនឹងជំងឺទឹកនោមផ្អែមនិងប្រឆាំងនឹងជំងឺមហារីក។ ទាំងនេះក៏ត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការថយចុះហានិភ័យនៃជំងឺមហារីកក្រពះនិងក្រពេញប្រូស្តាតផងដែរ។ ខ្ទឹមបារាំងនិងខ្ទឹមសត្រូវបានគេស្គាល់ថាមានសមាសធាតុ organosulfur ដែលជួយការពារការវិវត្តនៃជំងឺមហារីកដោយការបន្សាបជាតិពុលការថយចុះការលូតលាស់កោសិកាមហារីកនិងការរាំងស្ទះដល់ការឈឺទ្រូង។ ខ្ទឹមបារាំងនិងខ្ទឹមសក៏មានការប្រមូលផ្តុំខ្ពស់នៃសារជាតិប្រឆាំងអុកស៊ីតកម្ម flavonoid ដែលមានឥទ្ធិពលប្រឆាំងនឹងការរលាកដែលអាចជួយការពារជំងឺមហារីក។
ប៉េងប៉ោះ
ប៉េងប៉ោះសំបូរទៅដោយសារធាតុចិញ្ចឹមជាច្រើនប្រភេទដូចជាលីកូប៉ែនវីតាមីន C និងអ៊ីបេតាខារ៉ូទីននិងសារធាតុប្រឆាំងអុកស៊ីតកម្ម flavonol ។ លីកូប៉ែនអាចជួយការពារពីជំងឺមហារីកក្រពេញប្រូស្តាតការខូចស្បែកកាំរស្មីយូវីនិង? ជំងឺសរសៃឈាមបេះដូង។ លីកូប៉ែនត្រូវបានស្រូបយកបានល្អនៅពេលប៉េងប៉ោះត្រូវបានចម្អិន។ ទឹកប៉េងប៉ោះមួយពែងមានបរិមាណលីកូប៉ែនប្រហែល ១០ ដងនៃពែងប៉េងប៉ោះឆៅ។ សូមចងចាំផងដែរថា carotenoids ដូចជា lycopene ត្រូវបានស្រូបយកបានល្អបំផុតនៅពេលដែលអមដោយខ្លាញ់ដែលមានសុខភាពល្អដូច្នេះសូមរីករាយជាមួយប៉េងប៉ោះរបស់អ្នកនៅក្នុងសាឡាត់ជាមួយគ្រាប់ឬការស្លៀកពាក់ដែលមានមូលដ្ឋានលើសារធាតុចិញ្ចឹមដើម្បីទទួលបានអត្ថប្រយោជន៍អាហារូបត្ថម្ភបន្ថែម។
អាហារដែលយើងញ៉ាំអាចមានសក្តានុពលដែលអាចផ្តល់អត្ថប្រយោជន៍ឬបង្កគ្រោះថ្នាក់ដល់សុខភាពរបស់យើង។ អាហាររូបត្ថម្ភខ្សោយអាចបណ្តាលឱ្យមានបញ្ហាសុខភាពជាច្រើនរួមមានភាពធាត់ជំងឺបេះដូងនិងជំងឺទឹកនោមផ្អែមប្រភេទទី ២ ។ ទន្ទឹមនឹងនេះសារធាតុចិញ្ចឹមត្រឹមត្រូវអាចធ្វើឱ្យអ្នកមានអារម្មណ៍ស្វាហាប់កាត់បន្ថយហានិភ័យនៃបញ្ហាសុខភាពក៏ដូចជាជួយរក្សានិងរក្សាទម្ងន់ឱ្យមានសុខភាពល្អ។ ប្រសិនបើអ្នកចង់លើកកម្ពស់ភាពជាប់បានយូរអ្នកត្រូវដុតរាងកាយរបស់អ្នកដោយអាហារល្អ ៗ ។ អាហារល្អ ៗ ក៏អាចជួយកាត់បន្ថយការរលាកដែលទាក់ទងនឹងបញ្ហាសុខភាពផ្សេងៗគ្នារួមទាំងឈឺសន្លាក់និងរលាកសន្លាក់។ អ្នកជំនាញផ្នែកថែទាំសុខភាពដូចជាអ្នកជំនាញខាងជីវចេកអាចផ្តល់ការណែនាំអំពីរបបអាហារនិងរបៀបរស់នៅដើម្បីជួយលើកកម្ពស់សុខភាពនិងសុខភាព។ នៅក្នុងអត្ថបទបន្ទាប់យើងនឹងរាយបញ្ជីអាហារល្អ ៗ ជាច្រើនដែលទីបំផុតអាចជួយលើកកម្ពស់អាយុយឺនយូរ។ - វេជ្ជបណ្ឌិតអាឡិចហ្សង់ហ្សេនស៊ីស៊ីស៊ីធីស៊ីអ៊ិនធ័រ
Zesty Beet Juice
សេវាកម្ម: 1 ពេលចំអិន: 5-10 នាទី
ក្រូចថ្លុង ១ ចំណិតនិងចំណិត
ទឹកគ្រឿងផ្សំទាំងអស់នៅក្នុងទឹកផ្លែឈើដែលមានគុណភាពខ្ពស់។ បម្រើបានល្អបំផុតភ្លាមៗ។
គ្រាន់តែការ៉ុតមួយផ្តល់ឱ្យអ្នកនូវរាល់ការទទួលទានវីតាមីនអាប្រចាំថ្ងៃរបស់អ្នក
ត្រូវហើយការបរិភោគការ៉ុតតែ ៨០ ក្រាម (២ តាហ្ស៊ី) មួយផ្តល់ឱ្យអ្នកនូវការ៉ូទីនគ្រប់គ្រាន់សម្រាប់រាងកាយរបស់អ្នកដើម្បីផលិតនូវវីតាមីនអា ១.៤៨០ មីក្រូក្រាម (ចាំបាច់សម្រាប់ការបង្កើតកោសិកាស្បែកឡើងវិញ) ។ នោះគឺច្រើនជាងការទទួលទានប្រចាំថ្ងៃនៃវីតាមីនអាដែលត្រូវបានណែនាំនៅសហរដ្ឋអាមេរិកដែលមានប្រហែល ៩០០mcg ។ វាជាការល្អបំផុតក្នុងការញ៉ាំការ៉ុតដែលបានចម្អិនព្រោះនេះធ្វើឱ្យជញ្ជាំងកោសិកាទន់ដែលអនុញ្ញាតឱ្យស្រូបយកសារធាតុបេតាការ៉ូទីនបន្ថែមទៀត។ ការបន្ថែមអាហារដែលមានសុខភាពល្អទៅក្នុងរបបអាហាររបស់អ្នកគឺជាមធ្យោបាយដ៏ល្អមួយដើម្បីធ្វើឱ្យសុខភាពទូទៅរបស់អ្នកមានភាពប្រសើរឡើង។
វិសាលភាពនៃព័ត៌មានរបស់យើងត្រូវបានកំណត់ចំពោះរោគសាស្ត្រសាច់ដុំឆ្អឹងសាច់ដុំរាងកាយសុខភាពនិងបញ្ហាសុខភាពងាយរងគ្រោះនិង / ឬអត្ថបទវេជ្ជសាស្ត្រប្រធានបទនិងការពិភាក្សា។ យើងប្រើពិធីសារសុខភាពនិងសុខភាពដែលមានមុខងារដើម្បីព្យាបាលនិងគាំទ្រដល់ការរងរបួសឬការរំខាននៃប្រព័ន្ធសាច់ដុំ។ ប្រកាសប្រធានបទប្រធានបទនិងការយល់ដឹងរបស់យើងទាក់ទងនឹងបញ្ហាគ្លីនិកបញ្ហានិងប្រធានបទដែលទាក់ទងនិងគាំទ្រដោយផ្ទាល់ឬដោយប្រយោលនូវវិសាលភាពនៃការអនុវត្តគ្លីនិករបស់យើង។ ការិយាល័យរបស់យើងបានខិតខំប្រឹងប្រែងសមហេតុផលដើម្បីផ្តល់នូវឯកសារយោងដែលមានការគាំទ្រនិងបានកំណត់ការសិក្សាស្រាវជ្រាវដែលពាក់ព័ន្ធឬ សិក្សាគាំទ្រដល់ប្រកាសរបស់យើង។ យើងក៏ធ្វើការថតចម្លងការសិក្សាស្រាវជ្រាវដែលមានដល់ក្រុមប្រឹក្សាភិបាលឬសាធារណៈជនតាមការស្នើសុំ។ យើងយល់ថាយើងគ្របដណ្តប់លើបញ្ហានានាដែលត្រូវការការពន្យល់បន្ថែមអំពីវិធីដែលវាអាចជួយក្នុងផែនការថែទាំឬពិធីសារព្យាបាលជាក់លាក់។ ដូច្នេះដើម្បីពិភាក្សាបន្ថែមលើប្រធានបទខាងលើនេះសូមសួរលោកបណ្ឌិត Alex Jimenez ឬទាក់ទងមកយើងខ្ញុំតាម 915-850-0900
រៀបចំដោយវេជ្ជបណ្ឌិតអាឡិចជីមហ្សេនស៊ីស៊ីស៊ីធីស៊ី
ឯកសារយោង:
Joel Fuhrman, MD ។ ១០ អាហារល្អបំផុតដែលអ្នកអាចញ៉ាំដើម្បីរស់នៅបានយូរនិងមានសុខភាពល្អ ។ សុខភាពណាស់វែលវែន។ ថ្ងៃទី 6 ខែមិថុនា ឆ្នាំ 2020, www.verywellhealth.com/best-foods-for-longevity-4005852។
Dowden, Angela ។ កាហ្វេជាផ្លែឈើនិងជាអាហារពិតដែលមិនគួរឱ្យជឿ ។ របៀបរស់នៅ MSN ថ្ងៃទី 4 ខែមិថុនា ឆ្នាំ 2020, www.msn.com/en-us/foodanddrink/did-you-know/coffee-is-a-fruit-and-other-unbelievably-true-food-facts/ss-BB152Q5q?li=BBnb7Kz&ocid =mailsignout#image=24.
by វេជ្ជបណ្ឌិត Alex Jimenez ប្រឆាំងនឹងភាពចាស់ , ឱសថមុខងារ , សុខភាព , សុខភាពស្បែក , សុខុមាលភាព
តើអ្នកមានអារម្មណ៍ថា៖
ស្បែកក្រហមជាពិសេសនៅបាតដៃ? ស្បែករឺសក់ស្ងួតរឺក៏រលោង? ស្បែកមុនឬស្បែកមិនល្អ? ក្រចកខ្សោយ? អេឌីម៉ា?
ប្រសិនបើអ្នកកំពុងជួបប្រទះនឹងស្ថានភាពទាំងនេះបន្ទាប់មកកូលេស្តេរ៉ុលកូឡាជែនរបស់អ្នកអាចទាប។
នៅទីនោះមាន ជាការសិក្សាថ្មី អំពីរបៀបដែលកូឡាជែនអាចធ្វើអោយប្រសើរឡើងនូវសមាសភាពរាងកាយនៅពេលវាត្រូវបានផ្សំជាមួយនឹងលំហាត់ប្រចាំថ្ងៃ។ កូឡាជែននៅក្នុងរាងកាយមានសមាសធាតុអាស៊ីដអាមីណូតែមួយគត់ដែលដើរតួយ៉ាងសំខាន់ក្នុងកាយវិភាគសាស្ត្រនៃរាងកាយ។ ប្រូតេអ៊ីនកូឡាជែនគឺជាប្រភពប្រមូលផ្តុំនៃគ្លីសេរីនប្រូលីននិងអ៊ីដ្រូហ្សិនហើយនៅពេលវាត្រូវបានគេប្រៀបធៀបទៅនឹងប្រូតេអ៊ីនដែលមាននៅក្នុងរបបអាហារដទៃទៀតវាធ្វើឱ្យកូឡាជែនក្លាយជាជម្រើសជាក់ស្តែងមួយដែលមានសក្តានុពលជាប្រូតេអ៊ីនរចនាសម្ព័ន្ធ។
In ការសិក្សា 2015 ក្រុមអ្នកស្រាវជ្រាវបានបង្ហាញពីរបៀបដែលថ្នាំបំប៉នកូឡាជែនមានប្រសិទ្ធភាពអាចធ្វើអោយប្រសើរឡើងនូវសមាសធាតុរាងកាយនៅក្នុងបុរសសកម្ម។ លទ្ធផលបានបង្ហាញពីរបៀបដែលបុរសម្នាក់ៗចូលរួមក្នុងការបណ្តុះបណ្តាលទំងន់យ៉ាងហោចណាស់បីដងក្នុងមួយសប្តាហ៍ហើយត្រូវបំពេញបន្ថែមជាមួយនឹងថ្នាំកូឡាជែនយ៉ាងតិច ១៥ ក្រាមដើម្បីទទួលបានសុខភាពអតិបរមា។ ការវាយតម្លៃដែលតេស្តផ្តល់ជូនគឺតេស្តកម្លាំងការវិភាគជីវៈជីវសាស្ត្រនិងជីវសាច់ដុំ។ ការធ្វើតេស្តទាំងនេះធ្វើឱ្យប្រាកដថាបុរស ៗ មានដំណើរការល្អបន្ទាប់ពីទទួលយកថ្នាំបំប៉នកូឡាជែនហើយលទ្ធផលបង្ហាញថាតើម៉ាសរាងកាយរបស់ពួកគេមានការកើនឡើងនៃម៉ាសរាងកាយគ្មានខ្លាញ់។ ការសិក្សាមួយទៀតបានបង្ហាញពីរបៀបបំពេញបន្ថែមប្រូតេអ៊ីនកូឡាជែននៅពេលដែលវាត្រូវបានផ្សំជាមួយនឹងការបណ្តុះបណ្តាលភាពធន់ដែលអាចបង្កើនម៉ាសសាច់ដុំនិងកម្លាំងសាច់ដុំជាមួយមនុស្សវ័យចំណាស់ក៏ដូចជាមនុស្សដែលមានជំងឺសរសៃប្រសាទ។
លក្ខណៈសម្បត្តិមានប្រយោជន៍ជាមួយកូឡាជែន
មាន លក្ខណៈសម្បត្តិមានប្រយោជន៍ជាច្រើន ថ្នាំគ្រាប់កូឡាជែនដែលអាចផ្តល់ដល់រាងកាយនៅពេលដែលវាត្រូវបានគេប្រើប្រាស់។ មានកូឡាជែនកូឡាជែននិងជែលលីនដែលអាចជួយកែលម្អរចនាសម្ព័ន្ធស្បែករបស់មនុស្ស។ ទោះបីជាមិនមានការសិក្សាជាច្រើនអំពីថ្នាំបំប៉នកូឡាជែនក៏ដោយក៏មានការសន្យាដ៏ល្អសម្រាប់តំបន់នៅលើដងខ្លួន។ ពួកគេគឺជា:
ម៉ាសសាច់ដុំ ៖ ថ្នាំបំប៉នកូឡាជែននៅពេលផ្សំជាមួយនឹងការបណ្តុះបណ្តាលកម្លាំងអាចជួយបង្កើនសាច់ដុំនិងកម្លាំងនៅក្នុងខ្លួន។ជំងឺរលាកសន្លាក់ ៖ ថ្នាំបំប៉នកូឡាជែនអាចជួយមនុស្សដែលមានជំងឺរលាកសន្លាក់ឆ្អឹង។ ការសិក្សាបង្ហាញ នៅពេលដែលជំងឺរលាកសន្លាក់ឆ្អឹងបានលេបថ្នាំបំប៉នកូឡាជែនពួកគេបានរកឃើញការធ្លាក់ចុះយ៉ាងខ្លាំងនៃការឈឺចាប់ដែលពួកគេកំពុងជួបប្រទះ។ការបត់បែនស្បែក : នៅ ការសិក្សា 2014 វាបានបញ្ជាក់ថាស្ត្រីដែលបានប្រើថ្នាំបំប៉នកូឡាជែននិងបានបង្ហាញពីភាពប្រសើរឡើងនៃការបត់បែនស្បែក។ កូឡាជែនក៏អាចត្រូវបានប្រើក្នុងការព្យាបាលតាមបែបប្រធានបទដើម្បីជួយឱ្យស្បែករបស់មនុស្សមានភាពប្រសើរឡើងដោយកាត់បន្ថយស្នាមជ្រីវជ្រួញនិងស្នាមជ្រីវជ្រួញ។
អាហារបំប៉នកូឡាជែនមិនត្រឹមតែផ្តល់នូវគុណប្រយោជន៍ដែលមានប្រយោជន៍ដល់តំបន់ជាក់លាក់នៅលើដងខ្លួនប៉ុណ្ណោះទេប៉ុន្តែវាមានប្រភេទកូឡាជែន ៤ ប្រភេទសំខាន់ៗហើយតួនាទីរបស់វានៅក្នុងរាងកាយមនុស្សក៏ដូចជាមុខងាររបស់វាដែរ។
វាយ 1 ៖ ប្រភេទកូឡាជែនប្រភេទ ១ មានចំនួន ៩០ ភាគរយនៃកូឡាជែនរបស់រាងកាយនិងបង្កើតជាសរសៃក្រាស់ដែលផ្តល់រចនាសម្ព័ន្ធដល់ស្បែកឆ្អឹងជាលិកាភ្ជាប់និងធ្មេញដែលមាននៅក្នុងខ្លួន។វាយ 2 ៖ ប្រភេទកូឡាជែនប្រភេទទី ២ ត្រូវបានបង្កើតឡើងដោយសរសៃដែលមានរាងធូររលុងដែលមាននៅក្នុងឆ្អឹងខ្ចីដែលជួយទ្រទ្រង់សន្លាក់នៅក្នុងខ្លួន។វាយ 3 ៖ កូឡាជែនប្រភេទទី ៣ ជួយទ្រទ្រង់រចនាសម្ព័ន្ធសាច់ដុំសរីរាង្គនិងសរសៃឈាមដែលធ្វើឱ្យប្រាកដថារាងកាយដំណើរការបានត្រឹមត្រូវ។វាយ 4 ៖ ប្រភេទកូឡាជែនប្រភេទទី ៤ ត្រូវបានរកឃើញនៅស្រទាប់ស្បែកមនុស្សគ្រប់រូបនិងជួយដល់ការច្រោះនៅក្នុងខ្លួន។
ដោយសារកូឡាជែន ៤ ប្រភេទនេះមាននៅក្នុងខ្លួនវាចាំបាច់ត្រូវដឹងថាកូឡាជែនអាចថយចុះតាមធម្មជាតិតាមពេលវេលាតាមវ័យព្រោះរាងកាយនឹងផលិតនូវគុណភាពកូឡាជែនទាបជាងមុន។ សញ្ញាមួយដែលអាចមើលឃើញនៃការថយចុះកូឡាជែនគឺនៅពេលដែលស្បែកនៅលើខ្លួនមនុស្សប្រែជារឹងមាំនិងទន់ក៏ដូចជាធ្វើឱ្យឆ្អឹងខ្ចីចុះខ្សោយដោយសារតែភាពចាស់។
កត្តាដែលអាចបំផ្លាញខូឡាជេន
ទោះបីជាកូឡាជែនអាចថយចុះតាមធម្មជាតិក៏ដោយក៏កត្តាជាច្រើនអាចបំផ្លាញ collagens ដែលបង្កគ្រោះថ្នាក់ដល់ស្បែក។ កត្តាគ្រោះថ្នាក់អាចរួមមាន៖
ស្ករនិងការ៉ុត : ជាតិស្ករចម្រាញ់និងជាតិស្ករ អាចជ្រៀតជ្រែក ជាមួយនឹងសមត្ថភាពរបស់កូឡាជែនក្នុងការជួសជុលដោយខ្លួនឯងលើស្បែក។ ដូច្នេះតាមរយៈការកាត់បន្ថយការប្រើប្រាស់ជាតិស្ករនិងជាតិស្ករក្នុងរាងកាយវាអាចកាត់បន្ថយផលប៉ះពាល់នៃសរសៃឈាមវ៉ែនតម្រងនោមនិងការដាច់ជាលិកា។ពន្លឺព្រះអាទិត្យ ៈទោះបីជាការទទួលបានពន្លឺព្រះអាទិត្យគ្រប់គ្រាន់អាចជួយឱ្យមនុស្សម្នាក់រីករាយនៅថ្ងៃក៏ដោយប៉ុន្តែត្រូវបានប៉ះពាល់នឹងពន្លឺព្រះអាទិត្យក្នុងរយៈពេលយូរ អាចបណ្តាលឱ្យខូចខាត ទៅស្បែកនិងបំផ្លាញ peptides collagen ។ ផលប៉ះពាល់នៃពន្លឺព្រះអាទិត្យខ្លាំងពេកអាចបណ្តាលឱ្យស្បែកមានអាយុនិងបង្កើតភាពតានតឹងអុកស៊ីតកម្មនៅក្នុងខ្លួន។ការជក់បារី ៖ នៅពេលមនុស្សជក់បារីវាអាច កាត់បន្ថយផលិតកម្មកូឡាជែន នៅក្នុងរាងកាយបណ្តាលឱ្យរាងកាយមានស្នាមជ្រីវជ្រួញមុនអាយុហើយប្រសិនបើរាងកាយត្រូវបានរងរបួសដំណើរការនៃការជាសះស្បើយនឹងមានភាពយឺតយ៉ាវនិងអាចបណ្តាលឱ្យមានជំងឺនៅក្នុងខ្លួន។ជំងឺអូតូអ៊ុយមីន ៈជំងឺអូតូអ៊ុយមីនមួយចំនួនក៏អាចធ្វើឱ្យខូចខាតដល់ផលិតកម្មកូឡាជែនដែរ។
សន្និដ្ឋាន
កូឡាជែនមានសារៈសំខាន់ណាស់សម្រាប់រាងកាយព្រោះវាជួយឱ្យស្បែកទន់ភ្លន់និងរឹងមាំ។ តាមធម្មជាតិវានឹងថយចុះនៅពេលមនុស្សកាន់តែចាស់ដូច្នេះការប្រើថ្នាំបំប៉នកូឡាជែនអាចធានាថារាងកាយអាចដំណើរការបានត្រឹមត្រូវ។ នៅពេលកត្តាបង្កគ្រោះថ្នាក់ប៉ះពាល់ដល់រាងកាយពួកគេអាចបញ្ឈប់ឬបំផ្លាញផលិតកម្មកូឡាជែននិងពន្លឿនដំណើរការនៃស្នាមជ្រីវជ្រួញមុនអាយុពីការបង្កើតដែលធ្វើឱ្យមនុស្សម្នាក់មើលទៅចាស់ជាងពួកគេ។ ខ្លះ ផលិតផល អាចជួយដល់សកម្មភាពកោសិការបស់រាងកាយដោយផ្តល់នូវស្ថេរភាពល្អប្រសើរជាងមុនជីវសាស្ត្រនិងភាពងាយស្រួលក្នុងការរំលាយអាហារ។
វិសាលភាពនៃព័ត៌មានរបស់យើងត្រូវបានកំណត់ចំពោះបញ្ហាផ្នែកសាច់ដុំឆ្អឹងសាច់ដុំនិងបញ្ហាសុខភាពសរសៃប្រសាទឬអត្ថបទវេជ្ជសាស្ត្រដែលមានមុខងារប្រធានបទនិងការពិភាក្សា។ យើងប្រើពិធីសារសុខភាពដែលមានមុខងារដើម្បីព្យាបាលការរងរបួសឬការរំខាននៃប្រព័ន្ធសាច់ដុំ។ ការិយាល័យរបស់យើងបានខិតខំប្រឹងប្រែងសមហេតុផលដើម្បីផ្តល់នូវឯកសារយោងដែលមានការគាំទ្រនិងបានកំណត់ការសិក្សាស្រាវជ្រាវដែលពាក់ព័ន្ធឬការសិក្សាគាំទ្រដល់ប្រកាសរបស់យើង។ យើងក៏ធ្វើការថតចម្លងការសិក្សាស្រាវជ្រាវដែលមានដល់ក្រុមប្រឹក្សាភិបាលឬសាធារណៈជនតាមការស្នើសុំ។ ដើម្បីពិភាក្សាបន្ថែមលើប្រធានបទខាងលើនេះសូមសួរលោកបណ្ឌិត Alex Jimenez ឬទាក់ទងមកយើងខ្ញុំតាម 915-850-0900 .
ឯកសារយោង:
ក្រុមហ៊ុន Bosch, Ricardo, et al ។ Mechanismsនៃការថតរូបនិងកាត់ត Photocarcinogenesis និងយុទ្ធសាស្រ្ត Photoprotective ជាមួយPhytochemicals. សារធាតុប្រឆាំងអុកស៊ីតកម្ម (បាហ្សលប្រទេសស្វីស) , MDPI, ២៦ មីនា ២០១៥, www.ncbi.nlm.nih.gov/pmc/articles/PMC26/ ។
Danby, F William ។ ភាពទន់ខ្សោយនិងស្បែកដែលមានវ័យចំណាស់៖ ស្ករនិងគ្លីសេរីន គ្លីនិកផ្នែកសើស្បែក , បណ្ណាល័យវេជ្ជសាស្ត្រជាតិអាមេរិក, 2010, www.ncbi.nlm.nih.gov/pubmed/20620757 ។
ជេននីងខេរីរីអាន។ កូឡាជែន - តើវាគឺជាអ្វីហើយតើវាល្អសំរាប់អ្វី? Healthline , ថ្ងៃទី ៩ ខែកញ្ញាឆ្នាំ ២០១៦, www.healthline.com/ កង្វះអាហារូបត្ថម្ភ / កូឡាជែន។
ជុគលេវីវីច, មីកែល។ ការសិក្សាថ្មីបង្ហាញពីគុណប្រយោជន៍នៃថ្នាំកូឡាជែនសម្រាប់បង្កើនសមាសធាតុផ្សំរបស់រាងកាយរួមជាមួយការធ្វើលំហាត់ប្រាណ។ រចនាសម្រាប់សុខភាព , ថ្ងៃទី ៣១ ខែឧសភាឆ្នាំ ២០១៩, blog.designsforhealth.com/node/31 ។
Knuutinen, A, et al ។ ការជក់បារីជះឥទ្ធិពលដល់ការសំយោគកូឡាជែននិងការបំប្លែងម៉ាទ្រីសបន្ថែមលើស្បែកមនុស្ស ទិនានុប្បវត្តិរោគសើស្បែកអង់គ្លេស , បណ្ណាល័យវេជ្ជសាស្ត្រជាតិអាមេរិក, មេសា។ 2002, www.ncbi.nlm.nih.gov/pubmed/11966688 ។
Proksch, E, et al ។ ការបំពេញបន្ថែមដោយផ្ទាល់នូវថ្នាំកូឡាជែនជាក់លាក់មានប្រសិទ្ធិភាពជន៍លើសរីរវិទ្យាស្បែករបស់មនុស្ស៖ ការសិក្សាទ្វេ - ពិការដោយប្រើ placebo ។ ឱសថសាស្ត្រស្បែកនិងសរីរវិទ្យា , បណ្ណាល័យវេជ្ជសាស្ត្រជាតិអាមេរិក, 2014, www.ncbi.nlm.nih.gov/pubmed/23949208 ។
Schauss, Alexander G, et al ។ ផលប៉ះពាល់ពីទំងន់ម៉ូលេគុលទាបដែលមានជាតិប្រូតេអ៊ីនពីសាច់មាន់ដែលមានជាតិប្រូតេអ៊ីនពីរាងកាយដោយណូវែលជីវគីមីកូឡាជែនស្តីពីការធ្វើអោយប្រសើរឡើងនូវរោគសញ្ញាទាក់ទងនឹងជំងឺរលាកសន្លាក់ឆ្អឹង៖ ការធ្វើរោគវិនិច្ឆ័យដោយចៃដន្យខ្វាក់ភ្នែកទ្វេនិងការត្រួតពិនិត្យដោយប្រើថូ។ ទិនានុប្បវត្តិគីមីវិទ្យាកសិកម្មនិងអាហារ , បណ្ណាល័យវេជ្ជសាស្ត្រជាតិអាមេរិក, 25 មេសា។ 2012, www.ncbi.nlm.nih.gov/pubmed/22486722 ។
Zdzieblik, Denise, et al ។ ការបន្ថែមសារធាតុប៉េត្រូឡាទីនក្នុងការរួមផ្សំជាមួយនឹងការបណ្តុះបណ្តាលភាពធន់ធ្វើឱ្យសមាសធាតុរាងកាយកើនឡើងនិងបង្កើនកម្លាំងសាច់ដុំចំពោះបុរសវ័យចំណាស់ Sarcopenic៖ ការសាកល្បងត្រួតពិនិត្យដោយចៃដន្យ។ ទស្សនាវដ្តីអាហារូបត្ថម្ភអង់គ្លេស , សារព័ត៌មានសាកលវិទ្យាល័យខេមប្រ៊ីជថ្ងៃទី ២៨ ខែតុលាឆ្នាំ ២០១៥ គេហទំព័រ www.ncbi.nlm.nih.gov/pmc/articles/PMC28/ ។
តាមរយៈការជូនដំណឹងដល់បុគ្គលអំពីវិធីដែលសាកលវិទ្យាល័យជាតិវិទ្យាសាស្ត្រសុខាភិបាលផ្តល់ចំណេះដឹងសម្រាប់មនុស្សជំនាន់ក្រោយសាកលវិទ្យាល័យផ្តល់នូវវិជ្ជាជីវៈវេជ្ជសាស្ត្រជាច្រើនប្រភេទសម្រាប់មុខនាទីវេជ្ជសាស្ត្រដែលមានមុខងារ។
VIDEO
�
�
by វេជ្ជបណ្ឌិត Alex Jimenez ប្រឆាំងនឹងភាពចាស់ , ជំងឺស្ទះសរសៃពោះវៀន , ពោះវៀននិងពោះវៀន , សុខភាព , អាហាររូបត្ថម្ភ , សុខុមាលភាព
តើអ្នកមានអារម្មណ៍ថា៖
ដូចជាអ្នកត្រូវបានគេធ្វើរោគវិនិច្ឆ័យថាមានជំងឺ Celiac, រោគសញ្ញានៃការរលាកពោះវៀនធំ, Diverticulosis / Diverticulitis ឬរោគសញ្ញា Leaky Gut? ការក្រវាត់ក្រវាត់លំពែងហួសឬ ហើមពោះ? ការវង្វេងស្មារតីមិនធម្មតាបន្ទាប់ពីការព្យាបាលដោយប្រូតូលីកឬថ្នាំបំប៉នធម្មជាតិ? ការសង្ស័យនៃការទទួលទានអាហារបំប៉នមិនត្រឹមត្រូវ? តើបញ្ហារំលាយអាហារនៅស្ងៀមជាមួយនឹងការសំរាកលំហែទេ?
ប្រសិនបើអ្នកកំពុងជួបប្រទះនឹងស្ថានភាពទាំងនេះបន្ទាប់មកអ្នកប្រហែលជាកំពុងជួបប្រទះនឹងបញ្ហាវៀនហើយប្រហែលជាត្រូវសាកល្បងកម្មវិធី 4R ។
ភាពប្រែប្រួលនៃអាហារការរលាកសន្លាក់និងការថប់បារម្ភត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងភាពខ្សោយនៃក្រពះពោះវៀន។ លក្ខខណ្ឌផ្សេងៗទាំងនេះអាចកើតឡើងពីកត្តាជាច្រើនដែលអាចប៉ះពាល់ដល់បំពង់រំលាយអាហារ។ ប្រសិនបើទុកចោលមិនបានព្យាបាលវាអាចជាលទ្ធផលនៃភាពមិនដំណើរការនៃរបាំងភាពអសមត្ថភាពរបស់ពោះវៀនបណ្តាលឱ្យរលាកនិងស្ថានភាពសុខភាពធ្ងន់ធ្ងរដែលពោះវៀនអាចវិវត្តបាន។ កម្មវិធី 4R ត្រូវបានប្រើដើម្បីស្តារពោះវៀនដែលមានសុខភាពល្អនៅក្នុងខ្លួនហើយពាក់ព័ន្ធនឹងជំហានចំនួនបួន។ ពួកគេគឺ: ដកចេញជំនួសការបង្រួបបង្រួមនិងជួសជុល។
ការអនុញ្ញាតពីពោះវៀន។
ភាពអសមត្ថភាពរបស់ពោះវៀនជួយការពាររាងកាយនិងធានាថាបាក់តេរីដែលបង្កគ្រោះថ្នាក់មិនចូលក្នុងពោះវៀន។ វាការពាររាងកាយពី កត្តាបរិស្ថានដែលមានសក្តានុពល ដែលអាចបង្កគ្រោះថ្នាក់ហើយចូលក្នុងបំពង់រំលាយអាហារ។ វាអាចជាជាតិពុលមីក្រូសរីរាង្គបង្កជំងឺនិងពពួក Antigens ផ្សេងទៀតដែលអាចបង្កអន្តរាយដល់បំពង់រំលាយអាហារដែលបង្កបញ្ហា។ ស្រទាប់ខាងក្នុងនៃពោះវៀនមានស្រទាប់នៃកោសិកា epithelial ដែលត្រូវបានបំបែកដោយចំនុចប្រសព្វតឹង។ នៅក្នុងពោះវៀនដែលមានសុខភាពល្អ , ប្រសព្វតឹងរឹងធ្វើឱ្យអន្តរាយដល់ពោះវៀនដោយជ្រើសរើសសារធាតុចូលនិងធ្វើដំណើរឆ្លងកាត់របាំងពោះវៀននិងការពារកត្តាគ្រោះថ្នាក់ពីការស្រូបចូល។
កត្តាបរិស្ថានមួយចំនួនអាចធ្វើឱ្យខូចខាតដល់ប្រសព្វតឹងហើយលទ្ធផលគឺវាអាចបង្កើនភាពផុយស្រួយនៃពោះវៀនដែលបណ្តាលឱ្យមានភាពច្របូកច្របល់នៃពោះវៀនឬពោះវៀនលេចធ្លាយនៅក្នុងខ្លួន។ កត្តារួមចំណែកអាចបង្កើនភាពអសមត្ថភាពរបស់ពោះវៀនដូចជាបរិមាណខ្លាញ់និងអាល់កុលឆ្អែតច្រើនពេកកង្វះសារធាតុចិញ្ចឹមភាពតានតឹងរ៉ាំរ៉ៃនិងជំងឺឆ្លង។
ជាមួយនឹងការកើនឡើងនៃភាពជ្រាបនៃពោះវៀន នៅក្នុងពោះវៀនវាអាចធ្វើឱ្យអ្នកប្រឆាំងអុកស៊ីសែនឆ្លងទឹករំអិលពោះវៀនហើយចូលទៅក្នុងចរន្តឈាមដែលបណ្តាលឱ្យមានការឆ្លើយតបនៃប្រព័ន្ធភាពស៊ាំនិងការរលាកដល់រាងកាយ។ មានជម្ងឺក្រពះពោះវៀនមួយចំនួនដែលត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងភាពមិនធម្មតានៃពោះវៀនហើយប្រសិនបើមិនបានព្យាបាលវាអាចបង្កឱ្យមានស្ថានភាពស្វ័យប្រវត្ដិជាក់លាក់ដែលអាចបណ្តាលឱ្យមានគ្រោះថ្នាក់ដល់រាងកាយ។
កម្មវិធី 4Rs
4Rs គឺជាកម្មវិធីមួយដែលអ្នកជំនាញថែទាំសុខភាពណែនាំដល់អ្នកជំងឺរបស់ពួកគេឱ្យប្រើនៅពេលពួកគេកំពុងដោះស្រាយបញ្ហារំលាយអាហារដែលរំខាននិងជួយគាំទ្រដល់ការព្យាបាលពោះវៀន។
ការដោះបញ្ហា
ជំហានដំបូងនៅក្នុងកម្មវិធី 4Rs គឺត្រូវយកធាតុបង្កជំងឺដែលបង្កគ្រោះថ្នាក់និងការរលាកដែលត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការកើនឡើងនៃភាពធន់នឹងពោះវៀន។ កេះដូចជាស្ត្រេសនិងការផឹកស្រារ៉ាំរ៉ៃអាចបង្កអន្តរាយដល់រាងកាយរបស់មនុស្សម្នាក់ៗ។ ដូច្នេះការកំណត់គោលដៅកត្តាគ្រោះថ្នាក់ទាំងនេះចេញពីរាងកាយគឺត្រូវព្យាបាលវាដោយប្រើថ្នាំអង់ទីប៊ីយោទិចថ្នាំគ្រាប់និងការដកអាហារដែលរលាកចេញពីរបបអាហារត្រូវបានណែនាំរួមមាន៖
- អាល់កុល - Gluten - គ្រឿងបន្ថែមអាហារ - ផ្កាយ - អាស៊ីតខ្លាញ់ជាក់លាក់ - អាហារមួយចំនួនដែលមនុស្សម្នាក់ងាយនឹងញ៉ាំ
ការជំនួសសារធាតុចិញ្ចឹម
ជំហានទីពីរនៃកម្មវិធី 4Rs គឺដើម្បីជំនួសសារធាតុចិញ្ចឹមដែលបណ្តាលឱ្យមានបញ្ហាពោះវៀនតាមរយៈការរលាក។ សារធាតុចិញ្ចឹមជាក់លាក់អាចជួយកាត់បន្ថយការរលាកនៅក្នុងពោះវៀនខណៈពេលដែលធ្វើឱ្យប្រាកដថាបំពង់រំលាយអាហារត្រូវបានគាំទ្រ។ មានអាហារប្រឆាំងនឹងការរលាកមួយចំនួនដែលមានជីវជាតិ។ ទាំងនេះរួមបញ្ចូលទាំង:
- អាហារមានជាតិសរសៃខ្ពស់ - អូមេហ្គា -3s - ប្រេងអូលីវ - ផ្សិត - ឱសថប្រឆាំងនឹងការរលាក
មានថ្នាំបំប៉នជាក់លាក់មួយចំនួនអាចត្រូវបានប្រើដើម្បីទ្រទ្រង់មុខងាររំលាយអាហារដោយជួយនិងស្រូបយកសារធាតុចិញ្ចឹមដើម្បីជំរុញដល់សុខភាពពោះវៀន។ អ្វីដែលអង់ហ្ស៊ីមរំលាយអាហារធ្វើគឺពួកគេជួយក្នុងការបំបែកខ្លាញ់ប្រូតេអ៊ីននិងកាបូអ៊ីដ្រាតនៅក្នុងពោះវៀន។ នេះនឹងជួយដល់អ្នកដែលមានបញ្ហាប្រព័ន្ធរំលាយអាហារអន់ថយខ្វះអាហារឬមានជំងឺ celiac ។ ថ្នាំបំប៉នដូចជាថ្នាំគ្រាប់អាសុីតទឹកប្រមាត់អាចជួយក្នុងការស្រូបយកសារធាតុចិញ្ចឹមដោយការបញ្ចូលគ្នារវាងជាតិខ្លាញ់ក្នុងមាត់ជាមួយគ្នា។ ការសិក្សាបានបញ្ជាក់។ អាស៊ីតទឹកប្រមាត់នោះត្រូវបានប្រើដើម្បីព្យាបាលថ្លើមថង់ទឹកមាត់និងបំពង់ទឹកប្រមាត់ខណៈពេលដែលការពារការកកើតនៃគ្រួសក្នុងប្រមាត់បន្ទាប់ពីការវះកាត់ដុំពក។
Reinoculated The Gut
ជំហានទីបីគឺកម្មវិធី 4rs ដើម្បីបង្រួបបង្រួមអតិសុខុមប្រាណពោះវៀនជាមួយបាក់តេរីមានប្រយោជន៍ដើម្បីលើកកម្ពស់មុខងារពោះវៀនដែលមានសុខភាពល្អ។ ការសិក្សាត្រូវបានបង្ហាញ ថ្នាំគ្រាប់ probiotic ត្រូវបានគេប្រើដើម្បីធ្វើឱ្យប្រសើរឡើងនូវពោះវៀនដោយការស្តារបាក់តេរីមានប្រយោជន៍។ ជាមួយនឹងថ្នាំបំប៉នទាំងនេះពួកគេផ្តល់នូវភាពប្រសើរឡើងដល់ពោះវៀនដោយចាក់សារធាតុប្រឆាំងនឹងការរលាកចូលទៅក្នុងខ្លួនជួយទ្រទ្រង់ប្រព័ន្ធភាពស៊ាំផ្លាស់ប្តូរសមាសធាតុអតិសុខុមប្រាណរបស់រាងកាយនិងកាត់បន្ថយភាពអសមត្ថភាពរបស់ពោះវៀននៅក្នុងប្រព័ន្ធរំលាយអាហារ។
ចាប់តាំងពីការ ថ្នាំ probiotics ត្រូវបានរកឃើញ នៅក្នុងអាហារដែលមានជាតិ fermented និងត្រូវបានគេចាត់ទុកថាជាបណ្តោះអាសន្នចាប់តាំងពីពួកគេមិនស្ថិតស្ថេរនៅក្នុងបំពង់រំលាយអាហារនិងមានប្រយោជន៍។ គួរឱ្យភ្ញាក់ផ្អើលពួកគេនៅតែមានឥទ្ធិពលលើសុខភាពមនុស្សដោយសារតែឥទ្ធិពលនៃពោះវៀនដោយផលិតវីតាមីននិងសមាសធាតុប្រឆាំងនឹងអតិសុខុមប្រាណដូច្នេះផ្តល់នូវភាពចម្រុះនិងមុខងារពោះវៀន។
ការជួសជុលហ្គូត
ជំហានចុងក្រោយនៃកម្មវិធី 4Rs គឺការជួសជុលវៀន។ ជំហាននេះទាក់ទងនឹងការជួសជុលស្រទាប់ពោះវៀននៃពោះវៀនដែលមានសារធាតុចិញ្ចឹមនិងឱសថជាក់លាក់។ ឱសថនិងថ្នាំបំប៉នទាំងនេះអាចជួយកាត់បន្ថយភាពជ្រាបនិងការរលាកក្នុងរាងកាយ។ ឱសថនិងថ្នាំបំប៉នមួយចំនួនរួមមាន៖
- ប្រទាលកន្ទុយក្រពើ - ស្ករកៅស៊ូ Chios mastic - ឌីជីអេលអិល (អាជ្ញាប័ណ្ណខ្សោយ) - ឫស Marshmallow - L-glutamine - អូមេហ្គា -3s Polyphenols - វីតាមីនឌី - ស័ង្កសី
សន្និដ្ឋាន
ដោយសារកត្តាជាច្រើនអាចជះឥទ្ធិពលយ៉ាងធ្ងន់ធ្ងរដល់ប្រព័ន្ធរំលាយអាហារតាមរបៀបដែលមានគ្រោះថ្នាក់ហើយអាចជាអ្នកចូលរួមចំណែកដល់ស្ថានភាពសុខភាពមួយចំនួន។ គោលដៅសំខាន់នៃកម្មវិធី 4Rs គឺដើម្បីកាត់បន្ថយកត្តាទាំងនេះដែលធ្វើឱ្យប៉ះពាល់ដល់ពោះវៀននិងកាត់បន្ថយការរលាកនិងបង្កើនភាពអសមត្ថភាពរបស់ពោះវៀន។ នៅពេលដែលអ្នកជំងឺកំពុងត្រូវបានណែនាំអំពីកត្តាមានប្រយោជន៍ដែល 4Rs ផ្តល់ឱ្យវាអាចនាំឱ្យមានពោះវៀនដែលមានសុខភាពល្អនិងជាសះស្បើយ។ ខ្លះ ផលិតផល នៅទីនេះដើម្បីជួយគាំទ្រដល់ប្រព័ន្ធក្រពះពោះវៀនដោយគាំទ្រពោះវៀនធ្វើអោយប្រសើរឡើងនូវការរំលាយអាហារជាតិស្ករនិងផ្តោតលើអាស៊ីដអាមីណូដែលមានគោលបំណងគាំទ្រដល់ពោះវៀន។
វិសាលភាពនៃព័ត៌មានរបស់យើងត្រូវបានកំណត់ចំពោះបញ្ហាផ្នែកសាច់ដុំឆ្អឹងសាច់ដុំនិងបញ្ហាសុខភាពសរសៃប្រសាទឬអត្ថបទវេជ្ជសាស្ត្រដែលមានមុខងារប្រធានបទនិងការពិភាក្សា។ យើងប្រើពិធីសារសុខភាពដែលមានមុខងារដើម្បីព្យាបាលការរងរបួសឬការរំខាននៃប្រព័ន្ធសាច់ដុំ។ ការិយាល័យរបស់យើងបានខិតខំប្រឹងប្រែងសមហេតុផលដើម្បីផ្តល់នូវឯកសារយោងដែលមានការគាំទ្រនិងបានកំណត់ការសិក្សាស្រាវជ្រាវដែលពាក់ព័ន្ធឬការសិក្សាគាំទ្រដល់ប្រកាសរបស់យើង។ យើងក៏ធ្វើការថតចម្លងការសិក្សាស្រាវជ្រាវដែលមានដល់ក្រុមប្រឹក្សាភិបាលឬសាធារណៈជនតាមការស្នើសុំ។ ដើម្បីពិភាក្សាបន្ថែមលើប្រធានបទខាងលើនេះសូមសួរលោកបណ្ឌិត Alex Jimenez ឬទាក់ទងមកយើងខ្ញុំតាម 915-850-0900 .
ឯកសារយោង:
ដឺសាន់ស្យាសស្ត្រេនណី et al ។ គ្រាប់ចុចខ្វះអាហារូបត្ថម្ភសម្រាប់ម៉ូឌុលរបាំងពោះវៀន ព្រំដែននៅក្នុងប្រព័ន្ធភាពស៊ាំ , Frontiers Media SA, 7 ធ្នូ។ 2015, www.ncbi.nlm.nih.gov/pmc/articles/PMC4670985/ ។
Ianiro, Gianluca, et al ។ ថ្នាំបំប៉នអង់ស៊ីមបន្ថែមក្នុងជំងឺក្រពះពោះវៀន ។ ការរំលាយអាហារគ្រឿងញៀនបច្ចុប្បន្ន , អ្នកបោះពុម្ពវិទ្យាសាស្ត្រ Bentham, 2016, www.ncbi.nlm.nih.gov/pmc/articles/PMC4923703/ ។
Mu, Qinghui, et al ។ ឡាក់គីហ្គូតជាសញ្ញាគ្រោះថ្នាក់សម្រាប់ជំងឺអូតូអ៊ុយមីន ព្រំដែន , ជួរមុខ, 5 ឧសភា 2017, www.frontiersin.org/articles/10.3389/fimmu.2017.00598/full ។
រេហ្សក, សានណុន, et al ។ អាហារបំប៉នដែលជាប្រភពនៃរបបអាហាររបស់អង្គការផ្ទាល់ ព្រំដែននៅអតិសុខុមជីវវិទ្យា , Frontiers Media SA, 24 សីហា។ 2018, www.ncbi.nlm.nih.gov/pmc/articles/PMC6117398/ ។
Sander, Guy R. , et al ។ ការរអាក់រអួលនៃមុខងាររារាំងពោះវៀនដោយ Gliadin ពាក់ព័ន្ធនឹងការផ្លាស់ប្តូរការបញ្ចេញប្រូតេអ៊ីនអាប៉ូតូទីក។ សារព័ត៌មាន FEBS ចនវីលីននិងសុនអិលធីឌី ៨ សីហា ២០០៥ febs.onlinelibrary.wiley.com/doi/full/8/j.febslet.2005 ។
Sartor, R Balfour ។ ការព្យាបាលដោយការធ្វើចលនានៃមីក្រូហ្វីលីព្យូមក្នុងជំងឺរលាកពោះវៀនធំ៖ ថ្នាំអង់ទីប៊ីយោទិចប្រូស្តាតនិងបុព្វេ។ ផ្នែកប្រព័ន្ធរំលាយអាហារ , បណ្ណាល័យវេជ្ជសាស្ត្រជាតិអាមេរិក, ឧសភា 2004, www.ncbi.nlm.nih.gov/pubmed/15168372 ។
by វេជ្ជបណ្ឌិត Alex Jimenez ប្រឆាំងនឹងភាពចាស់ , ការឈឺទ្រូងរ៉ាំរ៉ៃ , ការឈឺចាប់រ៉ាំរ៉ៃ , របបអាហារ , ការតមអាហារ , ឱសថមុខងារ , សុខភាព , វេជ្ជសាស្ត្រទូទៅ , សុខុមាលភាពមុខងាររួមបញ្ចូលគ្នា , ឱសថសមាហរណកម្ម , ញ៉ាំបង្អែក , សុខភាពធម្មជាតិ , អាហាររូបត្ថម្ភ , ProLon , ឱសថ , ចុះទម្ងន់ , សុខុមាលភាព
ការឈឺចាប់រ៉ាំរ៉ៃគឺជាបញ្ហាសុខភាពទូទៅមួយដែលប៉ះពាល់ដល់មនុស្សជាច្រើននៅសហរដ្ឋអាមេរិក។ ខណៈពេលដែលលក្ខខណ្ឌវេជ្ជសាស្រ្តមួយចំនួនដូចជាជំងឺ fibromyalgia និងរោគសញ្ញាឈឺចាប់ myofascial អាចបណ្តាលឱ្យមានការឈឺចាប់រ៉ាំរ៉ៃវាក៏អាចវិវឌ្ឍន៍ដោយសារតែបញ្ហាសុខភាពផ្សេងទៀត។ ការសិក្សាស្រាវជ្រាវបានរកឃើញថាការរីករាលដាលនៃការរលាកគឺជាបុព្វហេតុចំបងនៃការឈឺចាប់រ៉ាំរ៉ៃ។ ការរលាកគឺជាយន្តការការពារធម្មជាតិចំពោះការរងរបួសជំងឺឬការឆ្លង។ ប៉ុន្តែប្រសិនបើដំណើរការរលាកនៅតែបន្តយូរពេកវាអាចក្លាយជាបញ្ហា។
ការរលាកបង្ហាញពីប្រព័ន្ធភាពស៊ាំដើម្បីជាសះស្បើយនិងជួសជុលជាលិកាដែលខូចខាតក៏ដូចជាដើម្បីការពារខ្លួនប្រឆាំងនឹងបាក់តេរីនិងវីរុស។ យ៉ាងណាមិញការរលាករ៉ាំរ៉ៃអាចបណ្តាលអោយមានបញ្ហាសុខភាពជាច្រើនរួមទាំងរោគសញ្ញានៃការឈឺចាប់រ៉ាំរ៉ៃផងដែរ។ ការផ្លាស់ប្តូររបៀបរស់នៅដែលមានសុខភាពល្អអាចជួយគ្រប់គ្រងការឈឺចាប់រ៉ាំរ៉ៃប៉ុន្តែមុនដំបូងសូមអោយយើងយល់ពីមូលហេតុទូទៅនៃការឈឺចាប់រ៉ាំរ៉ៃ។
តើការរលាកស្រួចស្រាលជាអ្វី?
ការរលាកស្រួចស្រាវតាមលំដាប់លំដោយកើតឡើងបន្ទាប់ពីមានរបួសឬអ្វីមួយដែលសាមញ្ញដូចជាឈឺបំពង់ក។ វាគឺជាការឆ្លើយតបបែបធម្មជាតិជាមួយនឹងផលប៉ះពាល់អវិជ្ជមានមានន័យថាវាធ្វើការនៅក្នុងស្រុកនៅក្នុងតំបន់ដែលមានបញ្ហាសុខភាព។ រោគសញ្ញាទូទៅនៃការរលាកស្រួចស្រាលរួមមានការហើមក្រហមការឈឺចាប់ការឈឺចាប់និងការបាត់បង់មុខងារដូចដែលបានលើកឡើងដោយបណ្ណាល័យឱសថជាតិ។ នៅពេលដែលការរលាកស្រួចស្រាវមានការរីកចម្រើននៃសរសៃឈាមដែលបណ្តាលអោយលំហូរឈាមកើនឡើងហើយកោសិកាគ្រាប់ឈាមសនៅក្នុងតំបន់ដែលរងរបួសជួយជំរុញការស្តារឡើងវិញ។
អំឡុងពេលមានការរលាកធ្ងន់ធ្ងរ, សមាសធាតុដែលហៅថា cytokines ត្រូវបានបញ្ចេញដោយជាលិកាដែលខូច។ cytokines ដើរតួជា "សញ្ញាអាសន្ន" ដែលនាំមកលើកោសិកាភាពស៊ាំរបស់រាងកាយខ្លួនមនុស្សក៏ដូចជាអ័រម៉ូននិងសារធាតុចិញ្ចឹមជាច្រើនដើម្បីជួសជុលបញ្ហាសុខភាព។ លើសពីនេះទៅទៀត, សារធាតុដូចអ័រម៉ូនដែលត្រូវបានគេស្គាល់ថា prostaglandins បង្កឱ្យកកឈាមដើម្បីព្យាបាលជាលិកាដែលខូចហើយទាំងនេះក៏អាចធ្វើអោយកម្តៅនិងការឈឺចាប់ដែលជាផ្នែកនៃដំណើរការរលាក។ នៅពេលការខូចខាតឬរបួសត្រូវបានជាសះស្បើយការរលាកអាចថយចុះ។
តើការរលាករ៉ាំរ៉ៃគឺជាអ្វី?
មិនដូចការរលាកស្រួចស្រាវការរលាករ៉ាំរ៉ៃមានឥទ្ធិពលយូរអង្វែង។ ការរលាករ៉ាំរ៉ៃត្រូវបានគេស្គាល់ផងដែរថាជាការរលាកជាប្រចាំបង្កជាការរលាកកំរិតទាបនៅទូទាំងរាងកាយរបស់មនុស្សដូចដែលបានបង្ហាញដោយការកើនឡើងនៃសញ្ញាប្រព័ន្ធភាពស៊ាំដែលមាននៅក្នុងឈាមនិងជាលិកាកោសិកា។ ការរលាករ៉ាំរ៉ៃក៏អាចបណ្តាលឱ្យមានជំងឺនិងជំងឺផ្សេងៗ។ កម្រិតនៃការរលាកជួនកាលអាចបណ្តាលឡើងទោះបីជាមិនមានការរងរបួសជំងឺឬការឆ្លងក៏ដោយក៏វាអាចធ្វើឱ្យប្រព័ន្ធភាពស៊ាំមានប្រតិកម្មដែរ។
ជាលទ្ធផលប្រព័ន្ធភាពស៊ាំរបស់មនុស្សអាចចាប់ផ្តើមវាយប្រហារកោសិកាជាលិកាឬសរីរាង្គដែលមានសុខភាពល្អ។ អ្នកស្រាវជ្រាវនៅតែព្យាយាមស្វែងយល់ពីផលវិបាកនៃការរលាករ៉ាំរ៉ៃនៅក្នុងខ្លួនមនុស្សនិងយន្ដការដែលពាក់ព័ន្ធនឹងដំណើរការការពារធម្មជាតិនេះ។ ឧទាហរណ៍ការរលាករ៉ាំរ៉ៃត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងបញ្ហាសុខភាពជាច្រើនដូចជាជំងឺបេះដូងនិងជំងឺដាច់សរសៃឈាមខួរក្បាល។
ទ្រឹស្តីមួយបានបង្ហាញថានៅពេលដែលការរលាកនៅតែមាននៅក្នុងសរសៃឈាមវាអាចជួយលើកកម្ពស់ការប្រមូលផ្តុំបន្ទះ។ យោងតាមសមាគមន៍បេះដូងអាមេរិច (American Heart Association) ឬ AHA បានអោយដឹងថាប្រសិនបើប្រព័ន្ធភាពស៊ាំកំណត់បន្ទះដូចអ្នកឈ្លានពានបរទេសនោះកោសិកាឈាមសអាចប៉ុនប៉ងបិទបាំងបន្ទះដែលរកឃើញនៅក្នុងឈាមហូរតាមសរសៃឈាម។ នេះអាចបង្កើតកំណកឈាមដែលអាចទប់ស្កាត់លំហូរឈាមទៅកាន់បេះដូងឬខួរក្បាលដែលបណ្តាលអោយវាក្លាយទៅជាគ្មានស្ថេរភាពនិងរលាក។ ជំងឺមហារីកគឺជាបញ្ហាសុខភាពមួយទៀតដែលត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការរលាករ៉ាំរ៉ៃ។ លើសពីនេះទៅទៀតយោងតាមវិទ្យាស្ថានជំងឺមហារីកជាតិការខូចខាត DNA អាចបណ្តាលមកពីការរលាករ៉ាំរ៉ៃផងដែរ។
ការរលាកដែលមិនធម្មតាជាប្រចាំមិនមានរោគសញ្ញាទេប៉ុន្តែអ្នកឯកទេសសុខាភិបាលអាចពិនិត្យរកប្រូតេអ៊ីន C-reactive ឬ CRP ដែលត្រូវបានគេស្គាល់ថាអាស៊ីត lipoic ដែលជាសញ្ញានៃការរលាកដែលត្រូវបានរកឃើញនៅក្នុងឈាម។ កម្រិតខ្ពស់នៃ CRP ត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការកើនឡើងហានិភ័យនៃជំងឺសរសៃឈាមបេះដូង។ កម្រិត CRP ខ្ពស់អាចត្រូវបានគេរកឃើញនៅក្នុងបញ្ហារ៉ាំរ៉ៃដូចជាជំងឺលុយពីសឬជំងឺរលាកសន្លាក់រ៉ាំរ៉ៃ។
ក្នុងករណីដែលមានជម្ងឺរ៉ាំរ៉ៃផ្សេងៗដូចជា fibromyalgia ប្រព័ន្ធសរសៃប្រសាទលើសពីប្រតិកម្មទៅនឹងការភ្ញោចជាក់លាក់វាគឺជាការរលាកដែលបណ្តាលអោយមានរោគសញ្ញានៃការឈឺចាប់រ៉ាំរ៉ៃ។ ប្រធានបទវាមិនអាចទៅរួចទេដែលអាចប្រាប់ពីភាពខុសគ្នារវាងការឈឺចាប់រ៉ាំរ៉ៃដែលបណ្តាលមកពីប្រព័ន្ធសរសៃប្រសាទដែលអន់ថយនិងការឈឺចាប់រ៉ាំរ៉ៃដែលបណ្តាលមកពីការរីករាលដាលនៃការរលាក។ ក្រៅពីការស្វែងរកតម្រុយនៅក្នុងចរន្តឈាមនោះអាហាររូបត្ថម្ភ, ទម្លាប់រស់នៅរបស់ជីវិតនិងការប៉ះពាល់ផ្នែកបរិស្ថានរបស់មនុស្សក៏អាចបង្កើនការរលាករ៉ាំរ៉ៃផងដែរ។
ការរលាកគឺជាយន្តការការពារធម្មជាតិរបស់ប្រព័ន្ធភាពស៊ាំប្រឆាំងនឹងការរងរបួសជំងឺឬការឆ្លង។ ខណៈពេលដែលការឆ្លើយតបរលាកនេះអាចជួយឱ្យជាសះស្បើយនិងជួសជុលជាលិកាការរលាករ៉ាំរ៉ៃអាចបណ្តាលឱ្យមានបញ្ហាសុខភាពជាច្រើនរួមទាំងរោគសញ្ញាឈឺចាប់រ៉ាំរ៉ៃ។ មានតុល្យភាព អាហាររូបត្ថម្ភរួមទាំងរបបអាហារផ្សេងៗនិងការតមអាហារអាចជួយកាត់បន្ថយការរលាក។ ការតមអាហារដែលត្រូវបានគេស្គាល់ផងដែរថាជាការកម្រិតកាឡូរីជួយជំរុញឱ្យមានជំងឺកោសិកានិងការស្ទុះងើបឡើងវិញនៃកោសិកា។ របបអាហារត្រាប់តាមការតមអាហារដែលជាផ្នែកមួយនៃផែនការរបបអាហារដែលមានអាយុយឺនយូរគឺជាកម្មវិធីតមអាហារដែល“ ល្បិច” រាងកាយរបស់មនុស្សទៅជារដ្ឋតមដើម្បីទទួលបានអត្ថប្រយោជន៍នៃការតមអាហារបែបប្រពៃណី។ មុនពេលធ្វើតាមរបបអាហារណាមួយដែលបានពិពណ៌នានៅក្នុងអត្ថបទនេះត្រូវប្រាកដថាពិគ្រោះជាមួយវេជ្ជបណ្ឌិត។
បណ្ឌិត Alex Jimenez DC, CCST Insight
អាហារូបត្ថម្ភរបបអាហារការតមអាហារនិងការឈឺចាប់រ៉ាំរ៉ៃ
របបអាហារប្រឆាំងនឹងការរលាកភាគច្រើនគឺបរិភោគផ្លែឈើស្រស់និងបន្លែត្រីនិងខ្លាញ់។ ផែនការរបបអាហារមេឌីទែរ៉ាណេតាមវិធីជាឧទាហរណ៍គឺរបបអាហារប្រឆាំងនឹងការរលាកដែលជំរុញឱ្យញ៉ាំគ្រាប់បរិមាណកម្រិតមធ្យមញ៉ាំសាច់តិចតួចនិងផឹកស្រា។ ផ្នែកអាហារប្រឆាំងនឹងការរលាកដូចជាអាស៊ីតខ្លាញ់អូមេហ្គា -3, ការពាររាងកាយរបស់មនុស្សប្រឆាំងនឹង d aម៉ាយ នាំមកដោយការរលាក។
របបអាហារប្រឆាំងនឹងការរលាកក៏ពាក់ព័ន្ធនឹងការឃ្លាតឆ្ងាយពីអាហារដែលអាចជំរុញការរលាក។ វាល្អប្រសើរក្នុងការបន្ថយបរិមាណអាហារដែលអ្នកញ៉ាំដែលមានជាតិខ្លាញ់ខ្ពស់និងជាតិខ្លាញ់ឆ្អែតដូចជាសាច់។ លើសពីនេះទៅទៀតរបបអាហារប្រឆាំងនឹងការរលាកបានរារាំងការប្រើប្រាស់កាបូអ៊ីដ្រាតចម្រាញ់និងអាហារដូចជានំបុ័ងនិងអង្ករ។ ទាំងនេះក៏ជំរុញឱ្យកាត់បន្ថយការប្រើប្រាស់នៃម្សៅនិងប្រេងដែលត្រូវបានគេផ្ទុកទៅដោយអាស៊ីតខ្លាញ់អូមេហ្គា -6 ដូចជាផ្កាឈូករ័ត្ន, ផ្កាកុលាប និង ប្រេងពោត។
ការតមអាហារឬកំហិតកាឡូរីត្រូវបានគេស្គាល់ថាជាយូរមកហើយដើម្បីកាត់បន្ថយភាពតានតឹងអុកស៊ីតកម្មនិងបន្ថយយន្ដការនៃវ័យចំណាស់នៅក្នុងសារពាង្គកាយផ្សេងៗ។ ផលប៉ះពាល់នៃការតមអាហារទាក់ទងនឹងការស្លាប់កោសិកាកម្មវិធីឬ apoptosis ប្រតិចារិកប្រសិទ្ធភាពថាមពលទូរស័ព្ទជីវហ្សុនហ្ស៊ីមហ្ស៊ីមូនហ្ស៊ីនយន្តការប្រឆាំងអុកស៊ីតកម្មនិងចង្វាក់ចរន្ត។ ការតមអាហារក៏រួមចំណែកផងដែរដល់ការធ្វើឱ្យសរីរាង្គផ្សិតដែលត្រូវបានគេស្គាល់ថាជាហ្សែនហ្វាហ្គីដែលហ្សែននៅក្នុងមេដំបូលត្រូវបានជំរុញឱ្យឆ្លងកាត់ apoptosis ដែលជម្រុញការស្តារឡើងវិញនៃថ្នាំពុល។
ការតមអាហារបណ្តោះអាសន្នអាចជួយអ្នកក្នុងការប្រយុទ្ធប្រឆាំងនឹងការរលាកធ្វើអោយប្រសើរឡើងនូវការរំលាយអាហារនិងបង្កើនអាយុយឺនយូរ។ រាងកាយមនុស្សត្រូវបានរចនាឡើងដើម្បីអាចរស់បានក្នុងរយៈពេលយូរដោយគ្មានអាហារ។ ការសិក្សាស្រាវជ្រាវបានបង្ហាញថាការតមអាហារបណ្តោះអាសន្នអាចមានការផ្លាស់ប្តូរជាវិជ្ជមាននៅក្នុងសមាសភាពទាំងមូលនៃអតិសុខុមប្រាណរបស់អ្នក។ លើសពីនេះទៅទៀតការតមអាហារបណ្តោះអាសន្នអាចកាត់បន្ថយភាពធន់ទ្រាំអាំងស៊ុយលីនខណៈពេលដែលបង្កើនការឆ្លើយតបរបស់ប្រព័ន្ធភាពស៊ាំ។ ទីបំផុតការតមអាហារបណ្តោះអាសន្នអាចជំរុញការផលិតសារធាតុដែលគេស្គាល់ថា? -hydroxybutyrate ដែលរារាំងផ្នែកនៃប្រព័ន្ធភាពស៊ាំដែលពាក់ព័ន្ធនឹងជម្ងឺរលាកក៏ដូចជាកាត់បន្ថយការផលិតផ្លាកសញ្ញារលាកដូចជាស៊ីតូកូសនិងប្រូតេអ៊ីន C ប្រតិកម្ម។ , ឬ CRP, ដូចដែលបានរៀបរាប់ខាងលើ។
ផែនការរបបអាហារដែលមានអាយុកាលដែលត្រូវបានបង្ហាញនៅក្នុងសៀវភៅដោយវេជ្ជបណ្ឌិត Valter Longo បានលុបបំបាត់ការប្រើប្រាស់អាហារដែលបានកែច្នៃដែលអាចបង្កឱ្យមានការរលាកនិងលើកកម្ពស់សុខុមាលភាពនិងភាពយូរអង្វែង។ កម្មវិធីរបបអាហារតែមួយនេះមិនដូចរបបអាហារបែបប្រពៃណីច្រើនបំផុតទេដែលមិនជួយសម្រកទម្ងន់។ ទោះបីជាអ្នកអាចមានការកាត់បន្ថយទម្ងន់ក៏ដោយការសង្កត់ធ្ងន់ទៅលើកម្មវិធីរបបអាហារដ៏ពិសេសនេះគឺការបរិភោគអាហារដែលមានសុខភាពល្អ។ ផែនការរបបអាហារមានអាយុវែងត្រូវបានបង្ហាញដើម្បីជួយធ្វើឱ្យកោសិកាបង្កើតឡើងវិញនូវស្នាមកោសិកាដើមកាត់បន្ថយជាតិខ្លាញ់ពោះនិងទប់ស្កាត់ការបាត់បង់ឆ្អឹងនិងសាច់ដុំដែលទាក់ទងនឹងអាយុព្រមទាំងបង្កើតភាពធន់ទ្រាំទៅនឹងការវិវត្តទៅរកជំងឺសរសៃឈាមបេះដូងជំងឺភ្លេចភ្លាំងជំងឺទឹកនោមផ្អែមនិងជំងឺមហារីក។
របបអាហារដែលធ្វើឱ្យមានល្បឿនលឿនឬ FMD អនុញ្ញាតឱ្យអ្នកទទួលបានអត្ថប្រយោជន៍នៃការតមអាហារតាមបែបប្រពៃណីដោយមិនចាំបាច់បង្អត់អាហាររបស់អ្នក។ ភាពខុសគ្នាសំខាន់នៃជំងឺរលាក FMD នោះគឺថាជំនួសឱ្យការលុបបំបាត់ទាំងស្រុងនូវអាហារទាំងអស់អស់រយៈពេលជាច្រើនថ្ងៃឬច្រើនសប្តាហ៍អ្នកគ្រាន់តែដាក់កម្រិតកាឡូរីរបស់អ្នកសម្រាប់រយៈពេលប្រាំថ្ងៃក្នុងមួយខែ។ FMD អាចត្រូវបានអនុវត្តម្តងក្នុងមួយខែដើម្បីជួយលើកកម្ពស់សុខភាពនិងសុខភាពទូទៅ។
ខណៈពេលដែលនរណាម្នាក់អាចតាមដានជំងឺរលាក FMD ដោយខ្លួនឯងផ្ទាល់ ProLon ការតមអាហារតាមរបៀបញ៉ាំអាហារផ្តល់នូវកម្មវិធីអាហារ 5 ថ្ងៃដែលត្រូវបានគេដាក់បញ្ចូលនិងដាក់ស្លាករៀងៗខ្លួនជារៀងរាល់ថ្ងៃដែលបម្រើអាហារដែលអ្នកត្រូវការសម្រាប់ជម្ងឺអេដស៍ក្នុងបរិមាណច្បាស់លាស់និងបន្សំ។ កម្មវិធីអាហារនេះត្រូវបានបង្កើតឡើងពីអាហារត្រៀមខ្លួនជាស្រេចដើម្បីបរិភោគឬងាយស្រួលរៀបចំដែលមានមូលដ្ឋានលើរុក្ខជាតិដែលមានដូចជាបារ៍ស៊ុបអាហារសម្រន់អាហារបំប៉នបន្ថែមភេសជ្ជៈនិងទឹកតែ។ មុនពេលចាប់ផ្ដើម ProLon តមអាហារត្រាប់តាមរបបអាហារកម្មវិធី ៥ ថ្ងៃ ឬការកែប្រែរបៀបរស់នៅណាដែលបានរៀបរាប់ខាងលើសូមប្រាកដថានិយាយទៅកាន់អ្នកឯកទេសថែទាំសុខភាពដើម្បីរកមើលការព្យាបាលឈឺរ៉ាំរ៉ៃណាមួយដែលសមស្របសម្រាប់អ្នក។
វិសាលភាពនៃព័ត៌មានរបស់យើងត្រូវបានកំណត់ចំពោះការព្យាបាលជំងឺឆ្អឹងខ្នង, បញ្ហាឆ្អឹងខ្នងនិងអត្ថបទឱសថប្រធានបទនិងការពិភាក្សា។ ដើម្បីពិភាក្សាបន្ថែមលើប្រធានបទខាងលើសូមកុំអោយលោកវេជ្ជបណ្ឌិត Alex Jimenez ទាក់ទងមកកាន់យើងដោយសេរី 915-850-0900 .
រៀបចំដោយវេជ្ជ
ការពិភាក្សាប្រធានបទបន្ថែម៖ ការឈឺឆ្អឹងខ្នងស្រួចស្រាវ
ឈឺខ្នង គឺជាបុព្វហេតុមួយក្នុងចំណោមបុព្វហេតុទូទៅនៃពិការភាពនិងខកខានថ្ងៃធ្វើការនៅទូទាំងពិភពលោក។ រោគសញ្ញានៃការឈឺខ្នងគឺជាមូលហេតុទូទៅទី ២ សម្រាប់ការមកការិយាល័យគ្រូពេទ្យដែលមានចំនួនច្រើនជាងការឆ្លងមេរោគលើផ្លូវដង្ហើមផ្នែកខាងលើ។ ប្រហែលជា ៨០ ភាគរយនៃចំនួនប្រជាជននឹងឈឺខ្នងយ៉ាងហោចណាស់ម្តងក្នុងមួយជីវិតរបស់ពួកគេ។ ឆ្អឹងខ្នងរបស់អ្នកគឺជារចនាសម្ព័ន្ធស្មុគស្មាញដែលបង្កើតឡើងដោយឆ្អឹងសន្លាក់សរសៃចងនិងសាច់ដុំក្នុងចំណោមជាលិការទន់ដទៃទៀត។ ការរងរបួសនិង / ឬលក្ខខណ្ឌកាន់តែធ្ងន់ធ្ងរដូចជា ស៊ីឌីសំបុក អាចបណា្ខលឱ្យមានការឈឺចាប់ឆ្អឹងខ្នង។ ការរងរបួសកីឡាឬគ្រោះថ្នាក់ចរាចររថយន្តជាញឹកញាប់គឺជាមូលហេតុនៃការឈឺចាប់ជាញឹកញាប់បំផុតទោះយ៉ាងណាពេលខ្លះចលនាសាមញ្ញបំផុតអាចមានលទ្ធផលឈឺចាប់។ ជាសំណាងល្អជម្រើសព្យាបាលជំនួសដូចជាការថែទាំឆ្អឹងខ្នងអាចជួយបន្ថយការឈឺចាប់តាមរយៈការប្រើឆ្អឹងខ្នងនិងការលៃលកដោយដៃដោយធ្វើអោយប្រសើរឡើងនូវការឈឺចាប់។
XYMOGEN s រូបមន្តវិជ្ជាជីវៈដាច់ដោយឡែកគឺអាចរកបានតាមរយៈជ្រើសរើសអ្នកជំនាញថែទាំសុខភាពដែលមានអាជ្ញាប័ណ្ណ។ ការលក់អនឡាញនិងការបញ្ចុះតម្លៃនៃរូបមន្ត XYMOGEN ត្រូវបានហាមឃាត់យ៉ាងតឹងរ៉ឹង។
ដោយមោទនភាព, វេជ្ជ ធ្វើឱ្យរូបមន្ត XYMOGEN អាចប្រើបានតែចំពោះអ្នកជំងឺដែលស្ថិតក្រោមការថែទាំរបស់យើង។
សូមទូរស័ព្ទទៅការិយាល័យរបស់យើងដើម្បីឱ្យពួកយើងផ្ដល់ការពិគ្រោះយោបល់ជាមួយវេជ្ជបណ្ឌិតដើម្បីចូលដំណើរការភ្លាមៗ។
ប្រសិនបើអ្នកជាអ្នកជំងឺ គ្លីនិកវេជ្ជសាស្ត្រនិង Chiropractic រងរបួស អ្នកអាចសួរអំពី XYMOGEN ដោយការហៅ 915-850-0900 .
ដើម្បីភាពងាយស្រួលនិងការពិនិត្យឡើងវិញរបស់អ្នក XYMOGEN ផលិតផលសូមពិនិត្យតំណខាងក្រោម។ *XYMOGEN-Catalog- ទាញយក
* គោលនយោបាយ XYMOGEN ខាងលើទាំងអស់នៅតែមានសុពលភាព។
***
by វេជ្ជបណ្ឌិត Alex Jimenez ប្រឆាំងនឹងភាពចាស់ , របបអាហារ , ការតមអាហារ , ឱសថមុខងារ , សុខភាព , វេជ្ជសាស្ត្រទូទៅ , សុខុមាលភាពមុខងាររួមបញ្ចូលគ្នា , ឱសថសមាហរណកម្ម , សុខភាពធម្មជាតិ , អាហាររូបត្ថម្ភ , ProLon , ថ្នាំគ្រាប់ , ចុះទម្ងន់ , សុខុមាលភាព
ការប្រកាន់ខ្ជាប់នូវរបបអាហារជាក់លាក់ដើម្បីរក្សាអាហាររូបត្ថម្ភឱ្យបានត្រឹមត្រូវជួនកាលអាចធ្វើឱ្យការស្ត្រេស។ ការកែប្រែរបៀបរស់នៅធម្មជាតិគឺជាគន្លឹះនៃការផ្លាស់ប្តូរទម្លាប់នៃការញ៉ាំរបស់អ្នកហើយនេះអាចជួយអ្នកឱ្យរស់នៅបានយូរនិងមានសុខភាពល្អ។ ផែនការរបបអាហារដែលមានអាយុកាលដែលបង្កើតឡើងដោយវេជ្ជបណ្ឌិត Valter Longo គឺជាជម្រើសនៃការណែនាំអំពីការទទួលទានដែលផ្តោតលើការផ្លាស់ប្តូរទម្លាប់ទទួលទានរបស់អ្នកដើម្បីទទួលបានសុខភាពនិងសុខភាពល្អ។
ច្បាប់នៃផែនការរបបជីវភាពធានារ៉ាប់រង
ដោយគ្រាន់តែធ្វើតាមគន្លឹះអាហារូបត្ថម្ភខាងក្រោមអ្នកអាចធ្វើកំណែទម្រង់របបអាហារបច្ចុប្បន្នរបស់អ្នកហើយចាប់ផ្ដើមទទួលទានអាហារដែលមានសុខភាពល្អដោយមិនចាំបាច់មានភាពតានតឹងនៃរបបអាហារបែបប្រពៃណី។ ផែនការរបបអាហារមានអាយុវែងលុបបំបាត់ការប្រើប្រាស់អាហារដែលកែច្នៃដែលអាចបង្កឱ្យមានបញ្ហាសុខភាពផ្សេងៗគ្នានិងបង្កើនការប្រើប្រាស់សារធាតុចិញ្ចឹមដែលជំរុញឱ្យមានអាយុវែង។ កម្មវិធីរបបអាហារតែមួយគត់នេះបានចែករំលែកលទ្ធផលនៃការសិក្សាស្រាវជ្រាវប្រហែល 25 ឆ្នាំទាំងអស់លើដំណោះស្រាយដ៏សាមញ្ញមួយដែលអាចជួយមនុស្សដែលមានបទពិសោធន៍សុខុមាលភាពទូទៅតាមរយៈអាហារបំប៉នត្រឹមត្រូវ។
ទោះយ៉ាងណាមិនដូចរបបអាហារប្រពៃណីភាគច្រើនទេរបបអាហារដែលមានអាយុវែងមិនលើកកម្ពស់ការសម្រកទម្ងន់នោះទេ។ ទោះបីជាអ្នកអាចជួបប្រទះការថយចុះទំងន់ក៏ដោយក៏ការសង្កត់ធ្ងន់នៃកម្មវិធីរបបអាហារពិសេសនេះគឺទៅលើការបរិភោគអាហារដែលមានសុខភាពល្អ។ ផែនការរបបអាហារយូរអង្វែងត្រូវបានបង្ហាញថាជួយឱ្យអ្នកធ្វើឱ្យកោសិកាថ្មីមានដំណើរការឡើងវិញស្រកទម្ងន់និងកាត់បន្ថយខ្លាញ់ក្បាលពោះការពារឆ្អឹងនិងសាច់ដុំដែលទាក់ទងនឹងអាយុបង្កើតភាពធន់នឹងការវិវត្តទៅជាជំងឺសរសៃឈាមបេះដូងជំងឺភ្លេចភ្លាំងជំងឺទឹកនោមផ្អែមនិងមហារីក។ ដូចជាពង្រីកអាយុយឺនយូរ។ ខាងក្រោមនេះយើងនឹងសង្ខេបនូវការណែនាំអំពីសារធាតុចិញ្ចឹមទូទៅបំផុតទាំង ៨ នៃផែនការរបបអាហារយូរអង្វែងដែលជាយថាហេតុអាចជួយឱ្យអាយុជីវិតរបស់អ្នកកាន់តែយូរនិងមានសុខភាពល្អ។
ផែនការរបបអាហារមានអាយុវែងគឺជាកម្មវិធីរបបអាហារតែមួយគត់ដែលត្រូវបានរចនាដោយលោកវេជ្ជបណ្ឌិត Valter Longo ដើម្បីលើកកម្ពស់សុខមាលភាពសុខភាពនិងអាយុជាយូរអង្វែង។ តាមរយៈការផ្លាស់ប្តូររបៀបរស់នៅសាមញ្ញមនុស្សអាចផ្លាស់ប្តូរទម្លាប់នៃការញ៉ាំរបស់ពួកគេនិងទាញយកប្រយោជន៍ពីអត្ថប្រយោជន៍សុខភាពជាច្រើននៃកម្មវិធីរបបអាហារនេះ។ ដោយធ្វើតាមរបបអាហារ pescatarian និងធ្វើតាម ProLon ការតមអាហារមីងឃីង ក្នុងចំណោមគន្លឹះអាហារូបត្ថម្ភដែលបានរៀបរាប់ខាងក្រោមមនុស្សអាចរស់នៅបានយូរនិងមានសុខភាពល្អ។ របបអាហារប្រពៃណីជាញឹកញាប់អាចមានភាពលំបាកនិងភាពតានតឹងក្នុងការធ្វើតាមប៉ុន្តែផែនការរបបអាហារយូរអង្វែងគឺជាកម្មវិធីរបបអាហារដែលមានប្រយោជន៍និងមានតែមួយគត់ដែលអាចមានលក្ខណៈសមរម្យសម្រាប់មនុស្សជាច្រើន។
បណ្ឌិត Alex Jimenez DC, CCST Insight គន្លឹះអាហារូបត្ថម្ភនៃរបបអាហារដែលមានអាយុវែង
អនុវត្តតាមរបបអាហារ Pescatarian
ក្នុងនាមជាផ្នែកមួយនៃផែនការរបបអាហាររយៈពេលវែងធ្វើតាមរបបអាហារ pescatarian ដែលស្ទើរតែ 100 ភាគរយនិងរុក្ខជាតិត្រី។ គួរប្រុងប្រយ័ត្នក្នុងការកំណត់ការប្រើត្រីដល់ 2 ឬ 3 ដងក្នុងមួយសប្តាហ៍ដោយជៀសវាងត្រីដែលមានជាតិបារតខ្ពស់ដូចជាត្រីធូណាត្រីដាវត្រីកណ្តុរនិងម្ទេស។ ប្រសិនបើអ្នកមានជាង 65 ហើយអ្នកចាប់ផ្តើមមានការថយចុះម៉ាសសាច់ដុំកម្លាំងនិងជាតិខ្លាញ់បន្ថែមត្រីបន្ថែមទៀតទៅក្នុងរបបអាហាររបស់អ្នកជាមួយនឹងចំណីអាហារដែលមានមូលដ្ឋានលើសត្វផ្សេងទៀតរួមទាំងស៊ុតនិងឈីសជាក់លាក់ដូចជា feta ឬ pecorino និងទឹកដោះគោដែលត្រូវបានធ្វើពីសត្វពពែ ទឹកដោះគោ។
កុំបរិគប្រូតេអ៊ីនច្រើនពេក
យោងទៅតាមផែនការរបបអាហារយូរអង្វែងយើងគួរញ៉ាំប្រូតេអ៊ីន 0.31 ទៅ 0.36 ក្រាមក្នុងមួយផោននៃជាតិខ្លាញ់ក្នុងរាងកាយជារៀងរាល់ថ្ងៃ។ ប្រសិនបើអ្នកថ្លឹងទម្ងន់ 130lbs អ្នកគួរតែញ៉ាំប្រូតេអ៊ីនពី 40 ទៅ 47 ក្រាម ថ្ងៃ, ឬសមមូលនៃត្រីសាម៉ុង 1.5, ពែងសឺរ 1 ឬសណ្តែកសណ្តែក 2 1 / 2 ដែលក្នុងនោះមាន 30 ក្រាមក្នុងមួយអាហារ។ ប្រសិនបើអ្នកមានទម្ងន់ចាប់ពី 200 ដល់ 220lbs, អ្នកគួរតែញ៉ាំប្រូតេអ៊ីនពី 60 ទៅ 70 ក្រាមក្នុងមួយថ្ងៃឬស្មើនឹងពីរសាឡាត់ត្រីសាឡី, 3 1 / 2 CUPS ឬ 1 1 / 2 CU of chickpeas ។ ការប្រើប្រាស់ប្រូតេអ៊ីនគួរតែត្រូវបានកើនឡើងបន្ទាប់ពីអាយុ 65 ។ សម្រាប់ភាគច្រើននៃពួកយើងការកើនឡើង 10 ទៅ 20 ភាគរយ, ឬ 5 ទៅ 10 ក្រាមច្រើនទៀតជារៀងរាល់ថ្ងៃ, គឺគ្រប់គ្រាន់។ ទីបំផុតរបបអាហារដែលមានអាយុកាលគឺមិនមានប្រូតេអ៊ីនសត្វដូចជាសាច់ក្រហមសាច់សត្វនិងបសុបក្សីលើកលែងតែប្រូតេអ៊ីនសត្វនៅក្នុងត្រី។ កម្មវិធីរបបអាហារដែលមានតែមួយគត់នេះគឺមានកំរិតខ្ពស់នៅក្នុងប្រូតេអ៊ីនបន្លែដូចជាគ្រាប់ពូជនិងគ្រាប់ដើម្បីបង្កើនសុខភាពនិងសុខភាពល្អ។
បង្កើនជាតិខ្លាញ់និងកាបូអ៊ីដ្រាតដ៏ស្មុគស្មាញ
ជាផ្នែកមួយនៃផែនការរបបអាហារដែលមានអាយុវែងអ្នកគួរតែញ៉ាំខ្លាញ់ច្រើនដែលមិនមានជាតិ polyunsaturated ដូចដែលគេរកឃើញនៅក្នុងត្រីសាម៉ុងអាល់ម៉ុនគ្រាប់វ៉ាល់ណាត់និងប្រេងអូលីវខណៈពេលដែលអ្នកគួរតែញ៉ាំជាតិខ្លាញ់ឆ្អែតអុកស៊ីដុលនិងអេត្យូពីស៊ី។ ដូចគ្នានេះដែរដែលជាផ្នែកមួយនៃផែនការរបបអាហារយូរអង្វែងអ្នកក៏គួរតែបរិភោគកាបូអ៊ីដ្រាតស្មុគស្មាញដូចជាអ្វីដែលត្រូវបានគេរកឃើញនៅក្នុងនំបុ័ងស្រូវសាលីធញ្ញជាតិនិងបន្លែ។ ត្រូវប្រាកដថាកំណត់ការញ៉ាំចំណីសត្វអង្ករនំប៉័ងផ្លែឈើនិងទឹកផ្លែឈើដែលអាចប្តូរទៅជាជាតិស្ករនៅពេលដែលវាឈានដល់វៀន។
ទទួលយកអាហារបំប៉ន
រាងកាយមនុស្សត្រូវការប្រូតេអ៊ីនអាស៊ីតខ្លាញ់សំខាន់ដូចជាអូមេហ្គា -3 និងអូមេហ្គា-6 វីតាមីនសារធាតុរ៉ែនិងស្ករសូម្បីតែដំណើរការឱ្យបានត្រឹមត្រូវ។ នៅពេលដែលការទទួលទានអាហារបំប៉នជាក់លាក់របស់អ្នកទាបពេកការជួសជុលការជំនួសនិងការការពាររបស់រាងកាយមនុស្សអាចបន្ថយល្បឿនឬបញ្ឈប់ដែលអាចឱ្យផ្សិតបាក់តេរីនិងវីរុសបង្កការខូចខាតដែលអាចនាំឱ្យមានបញ្ហាសុខភាពជាច្រើន។ ទទួលទានអាហារបំប៉នវីតាមីននិងសារធាតុរ៉ែជាពិសេសសម្រាប់អូមេហ្គា-3 តាមការណែនាំដោយអ្នកជំនាញថែទាំសុខភាពរបស់អ្នក។
ទទួលទានអាហារផ្សេងៗពីកncestry
ដើម្បីទទួលបានសារធាតុចិញ្ចឹមទាំងអស់ដែលអ្នកត្រូវការអ្នកត្រូវញ៉ាំចំណីអាហារជាច្រើនប៉ុន្តែវាជាការល្អបំផុតដែលអ្នកជ្រើសរើសអាហារដែលមាននៅលើតុឪពុកម្តាយជីដូនជីតានិងជីដូនជីតា។ ជាឧទាហរណ៍នៅប្រទេសអឺរ៉ុបភាគខាងជើងជាច្រើនដែលទឹកដោះគោត្រូវបានគេប្រើជាទូទៅការមិនអត់ធ្មត់ចំពោះ lactose គឺកម្រណាស់ប៉ុន្តែការមិនអត់ធ្មត់ចំពោះ lactose គឺមានជាទូទៅនៅក្នុងបណ្តាប្រទេសនៅអឺរ៉ុបភាគខាងត្បូងនិងនៅអាស៊ីដែលទឹកដោះគោមិនមែនជាផ្នែកមួយនៃរបបអាហារធម្មតារបស់មនុស្សពេញវ័យ។ ប្រសិនបើមនុស្សដែលមានដើមកំណើតជប៉ុនរស់នៅក្នុងសហរដ្ឋអាមេរិចភ្លាមៗសម្រេចចិត្តចាប់ផ្តើមផឹកទឹកដោះគោដែលកម្រនឹងបម្រើនៅតុបរិភោគអាហាររបស់ជីដូនជីតារបស់ពួកគេនោះពួកគេប្រហែលជាចាប់ផ្តើមឈឺ។ បញ្ហាទូទៅបំផុតក្នុងករណីទាំងនេះគឺការមិនអត់ឱនឬការទប់ស្កាត់ស្វ័យប្រវត្តដូចជាការឆ្លើយតបចំពោះអាហារដែលសម្បូរទៅដោយជាតិស្ករដូចជានំបុ័ងនិងប៉ាស្តាដែលត្រូវបានគេឃើញនៅក្នុងមនុស្សដែលមានជំងឺ celiac ។ ទោះបីជាត្រូវការអំណះអំណាងបន្ថែមក៏ដោយក៏វាអាចទៅរួចដែលថាការមិនអត់ធ្មត់ចំពោះអាហារអាចទាក់ទងទៅនឹងជំងឺអូតូអ៊ុយមីនជាច្រើនដែលរួមមានជំងឺទឹកនោមផ្អែមជំងឺរលាកពោះវៀនធំនិងជំងឺ Crohn ។
ទទួលទានអាហារពីរក្នុងមួយថ្ងៃនិងអាហារសម្រន់
យោងទៅតាមផែនការរបបអាហារយូរអង្វែងវាជាការប្រសើរក្នុងការទទួលទានអាហារពេលព្រឹកនិងអាហារសំខាន់មួយបូករួមនឹងអាហារសម្រន់ដែលមានកាឡូរីទាបនិងស្ករទាបជារៀងរាល់ថ្ងៃ។ ខណៈពេលដែលសម្រាប់មនុស្សមួយចំនួនវាអាចត្រូវបានគេណែនាំឱ្យញ៉ាំអាហារបីពេលនិងអាហារសម្រន់ជារៀងរាល់ថ្ងៃ។ គោលការណ៍ណែនាំអំពីសារធាតុចិញ្ចឹមជាច្រើនណែនាំថាយើងគួរទទួលទានអាហារពី ៥ ទៅ ៦ ពេលក្នុងមួយថ្ងៃ។ នៅពេលដែលមនុស្សត្រូវបានគេណែនាំឱ្យញ៉ាំជាញឹកញាប់វាអាចជាការពិបាកសម្រាប់ពួកគេក្នុងការគ្រប់គ្រងការទទួលទានកាឡូរី។ ក្នុងរយៈពេល ២០ ឆ្នាំចុងក្រោយនេះប្រជាជនប្រមាណ ៧០ ភាគរយនៃប្រជាជននៅសហរដ្ឋអាមេរិកត្រូវបានគេចាត់ទុកថាធាត់លើសទម្ងន់។ វាកាន់តែពិបាកក្នុងការទទួលទានអាហារបំប៉នអាយុវែងប្រសិនបើអ្នកញ៉ាំតែ ២ កន្លះកន្លះក្នុងមួយថ្ងៃ។ វានឹងត្រូវការចំណែកដ៏ច្រើននៃបន្លែផ្លែឈើនិងត្រីដើម្បីឈានដល់បរិមាណដែលនាំឱ្យឡើងទម្ងន់។ អាហារបំប៉នខ្ពស់បូករួមនឹងបរិមាណអាហារផ្ញើសញ្ញាទៅក្រពះនិងខួរក្បាលរបស់អ្នកថាអ្នកមានអាហារគ្រប់គ្រាន់។ ប្រព័ន្ធអាហារដ៏សំខាន់មួយនេះជួនកាលត្រូវបំបែកជាពីរអាហារដើម្បីចៀសវាងបញ្ហារំលាយអាហារ។ មនុស្សពេញវ័យនិងមនុស្សចាស់ងាយនឹងស្រកទំងន់គួរតែញ៉ាំអាហារបីពេលក្នុងមួយថ្ងៃ។ ចំពោះអ្នកដែលព្យាយាមបញ្ចុះទម្ងន់ក៏ដូចជាមនុស្សដែលធាត់លើសទម្ងន់ការណែនាំអាហារបំប៉នល្អបំផុតគឺទទួលទានអាហារពេលព្រឹកជារៀងរាល់ថ្ងៃ។ មានអាហារពេលល្ងាចឬអាហារថ្ងៃត្រង់ប៉ុន្តែមិនទាំងពីរហើយជំនួសឱ្យអាហារដែលខកខានដោយអាហារសម្រន់មួយដែលមានកាឡូរីតិចជាង ១០០ និងមិនលើសពី ៣ ទៅ ៥ ក្រាមនៃជាតិស្ករ។ តើអាហារណាដែលអ្នករំលងគឺអាស្រ័យលើរបៀបរស់នៅរបស់អ្នកទោះជាយ៉ាងណាក៏ដោយវាមិនត្រូវបានណែនាំឱ្យរំលងអាហារពេលព្រឹកដោយសារតែបញ្ហាសុខភាពមិនល្អ។ អត្ថប្រយោជន៍នៃការរំលងអាហារថ្ងៃត្រង់គឺពេលវេលានិងថាមពលឥតគិតថ្លៃ។ ប៉ុន្តែវាមានគុណវិបត្តិសម្រាប់ការបរិភោគអាហារពេលល្ងាចដ៏ធំជាពិសេសអ្នកដែលទទួលរងពីការចាល់អាស៊ីតឬបញ្ហាដំណេក។ គុណវិបត្តិសម្រាប់ការរំលងអាហារពេលល្ងាចទោះយ៉ាងណាវាអាចលុបបំបាត់អាហារសង្គមនៅសម័យរបស់ពួកគេ។
បរិភោគក្នុងរយៈពេល 12-Hour Window រាល់ថ្ងៃ
ទម្លាប់នៃការញ៉ាំជាទូទៅមួយទៀតដែលត្រូវបានអនុម័តដោយមនុស្សរាប់រយនាក់គឺជាការហាមឃាត់ការញ៉ាំអាហារឬអាហារសម្រន់គ្រប់ពេលក្នុងមួយថ្ងៃរៀងរាល់ថ្ងៃ។ ប្រសិទ្ធភាពនៃវិធីសាស្រ្តនេះត្រូវបានបង្ហាញនៅក្នុងការសិក្សាស្រាវជ្រាវទាំងមនុស្សនិងសត្វ។ ជាទូទៅអ្នកនឹងញ៉ាំអាហារពេលព្រឹកនៅម៉ោង 12 ហើយបន្ទាប់មកញ៉ាំអាហារពេលល្ងាចត្រឹមម៉ោង 20 រសៀល។ ការញ៉ាំចំណីអាហារមានរយៈពេល 10 ម៉ោងឬតិចជាងនេះអាចជាការប្រសើរជាងចំពោះការសម្រកទម្ងន់ប៉ុន្តែវាពិបាកក្នុងការរក្សាវាហើយវាអាចបង្កើនហានិភ័យនៃការវិវត្តន៍នៃផលប៉ះពាល់ដូចជាគ្រួសក្នុងថង់ទឹកប្រមាត់និងអាចបង្កើនឱកាសនៃការវិវត្តទៅជាជំងឺសរសៃឈាមបេះដូង។ អ្នកមិនគួរញ៉ាំ 3 ទៅ 4 ម៉ោងមុនពេលគេង។
សូមធ្វើតាមរបបអាហារProLon ការតមអាហារមីងឃីង
មនុស្សដែលមានសុខភាពល្អដែលមានអាយុក្រោម 65 គួរតែធ្វើតាម ProLon ការតមអាហារមីងឃីតកម្មវិធីអាហារ ៥ ថ្ងៃ យ៉ាងហោចណាស់ពីរដងជារៀងរាល់ឆ្នាំ។ ជំងឺនេះគឺជាគោលការណ៍សំខាន់មួយដែលត្រូវបានលើកកំពស់ដោយផែនការអាយុជីវិត។ របបអាហារដែលធ្វើត្រាប់តាមការញ៉ាំផ្តល់នូវអត្ថប្រយោជន៍សុខភាពដូចគ្នានៃការតមអាហារដោយមិនចាំបាច់តមអាហារ។ ដោយការញ៉ាំកាឡូរីពី 800 ទៅ 1,100 នៅក្នុងបរិមាណច្បាស់លាស់និងការរួមបញ្ចូលគ្នានៃអាហារដែលត្រូវបានគេដាក់និងដាក់ស្លាករៀងៗខ្លួនក្នុងមួយថ្ងៃអ្នកអាច "បោកបញ្ឆោត" រាងកាយមនុស្សក្នុងស្ថានភាពតមអាហារ។ តាមរយៈការស្រាវជ្រាវជាច្រើនវេជ្ជបណ្ឌិត Valter Longo បានរកឃើញថាដោយការបោសសំអាតរាងកាយនៃអាហារតាមរបៀបនេះកោសិការបស់យើងចាប់ផ្តើមបំបែកនិងបង្កើតជាលិកាខាងក្នុងរបស់យើងតាមរយៈដំណើរការដែលត្រូវបានគេស្គាល់ថា autophagy ការសំលាប់និងការជំនួសឬការបង្កើតឡើងវិញនូវកោសិកាដែលខូច។ លើសពីនេះទៀតការតមអាហារអាចផ្លាស់ប្តូរបញ្ហាសុខភាពជាច្រើន, បំផ្លាញកោសិកាមហារីកនិងកាត់បន្ថយលទ្ធភាពនៃការវិវត្តទៅជាជំងឺអាល់ហ្សៃមឺរ។
របស់អ្នក អាយុវែង។ នេះ វិសាលភាពនៃពត៌មានរបស់យើងត្រូវបានកំណត់ចំពោះបញ្ហាសុខភាពខួរក្បាលបញ្ហាឆ្អឹងខ្នងនិងប្រធានបទឱសថមុខងារ។ ដើម្បីពិភាក្សាបន្ថែមទៀតអំពីបញ្ហាប្រធានបទសូមមានអារម្មណ៍ស៊ប់ដោយសេរីដើម្បីសួរវេជ្ជបណ្ឌិតអេលជីមហ្ស៊ីលឺរឬទាក់ទងយើងនៅ 915-850-0900 .
រៀបចំដោយវេជ្ជ
ការពិភាក្សាប្រធានបទបន្ថែម៖ ការឈឺឆ្អឹងខ្នងស្រួចស្រាវ
ឈឺខ្នង គឺជាបុព្វហេតុមួយក្នុងចំណោមបុព្វហេតុទូទៅនៃពិការភាពនិងខកខានថ្ងៃធ្វើការនៅទូទាំងពិភពលោក។ រោគសញ្ញានៃការឈឺខ្នងគឺជាមូលហេតុទូទៅទី ២ សម្រាប់ការមកការិយាល័យគ្រូពេទ្យដែលមានចំនួនច្រើនជាងការឆ្លងមេរោគលើផ្លូវដង្ហើមផ្នែកខាងលើ។ ប្រហែលជា ៨០ ភាគរយនៃចំនួនប្រជាជននឹងឈឺខ្នងយ៉ាងហោចណាស់ម្តងក្នុងមួយជីវិតរបស់ពួកគេ។ ឆ្អឹងខ្នងរបស់អ្នកគឺជារចនាសម្ព័ន្ធស្មុគស្មាញដែលបង្កើតឡើងដោយឆ្អឹងសន្លាក់សរសៃចងនិងសាច់ដុំក្នុងចំណោមជាលិការទន់ដទៃទៀត។ ការរងរបួសនិង / ឬលក្ខខណ្ឌកាន់តែធ្ងន់ធ្ងរដូចជា ស៊ីឌីសំបុក អាចបណា្ខលឱ្យមានការឈឺចាប់ឆ្អឹងខ្នង។ ការរងរបួសកីឡាឬគ្រោះថ្នាក់ចរាចររថយន្តជាញឹកញាប់គឺជាមូលហេតុនៃការឈឺចាប់ជាញឹកញាប់បំផុតទោះយ៉ាងណាពេលខ្លះចលនាសាមញ្ញបំផុតអាចមានលទ្ធផលឈឺចាប់។ ជាសំណាងល្អជម្រើសព្យាបាលជំនួសដូចជាការថែទាំឆ្អឹងខ្នងអាចជួយបន្ថយការឈឺចាប់តាមរយៈការប្រើឆ្អឹងខ្នងនិងការលៃលកដោយដៃដោយធ្វើអោយប្រសើរឡើងនូវការឈឺចាប់។
XYMOGEN s រូបមន្តវិជ្ជាជីវៈដាច់ដោយឡែកគឺអាចរកបានតាមរយៈជ្រើសរើសអ្នកជំនាញថែទាំសុខភាពដែលមានអាជ្ញាប័ណ្ណ។ ការលក់អនឡាញនិងការបញ្ចុះតម្លៃនៃរូបមន្ត XYMOGEN ត្រូវបានហាមឃាត់យ៉ាងតឹងរ៉ឹង។
ដោយមោទនភាព, វេជ្ជ ធ្វើឱ្យរូបមន្ត XYMOGEN អាចប្រើបានតែចំពោះអ្នកជំងឺដែលស្ថិតក្រោមការថែទាំរបស់យើង។
សូមទូរស័ព្ទទៅការិយាល័យរបស់យើងដើម្បីឱ្យពួកយើងផ្ដល់ការពិគ្រោះយោបល់ជាមួយវេជ្ជបណ្ឌិតដើម្បីចូលដំណើរការភ្លាមៗ។
ប្រសិនបើអ្នកជាអ្នកជំងឺ គ្លីនិកវេជ្ជសាស្ត្រនិង Chiropractic រងរបួស អ្នកអាចសួរអំពី XYMOGEN ដោយការហៅ 915-850-0900 .
ដើម្បីភាពងាយស្រួលនិងការពិនិត្យឡើងវិញរបស់អ្នក XYMOGEN ផលិតផលសូមពិនិត្យតំណខាងក្រោម។ *XYMOGEN-Catalog- ទាញយក
* គោលនយោបាយ XYMOGEN ខាងលើទាំងអស់នៅតែមានសុពលភាព។
***